Iron in Initiation and Promotion of Human Cancer -How Free Iron Accelerates Predisposing Insulin Resistance
Juan Ariel Jara Guerrero1*
1Doctoral student in Medicine Complutense University of Madrid Department of Experimental Endocrinology, Spain.
*Corresponding Author:Juan Ariel Jara Guerrero, Doctoral student in Medicine Complutense University of MadridDepartment of Experimental Endocrinology,Spain, Tel: +34 913 94 13 07; Fax: +34 913 94 13 07
Citation: Juan Ariel Jara Guerrero (2023)Iron in Initiation and Promotion of Human Cancer -How Free Iron Accelerates Predisposing Insulin Resistance. Cancer Prog Diagn 7: 143.
Received: April 28, 2023; Accepted: May 28, 2023; Published: May 31, 2023.
Messages not to Forget
1) Iron overload in our diet generates Tissue and cellular Resistance to Insulin (93).
2) By reducing iron stores by donating blood, insulin sensitivity is increased, the early onset of Chronic-Degenerative Diseases is prevented, and Diabetes Mellitus in particular, even among people with normal serum Ferritin levels (Facchini, 2001; Fernández Real; 2005) (31, 107).
3) It has been repeatedly demonstrated, for more than 25 years, that the Malignant cell transformation induced by oxidative stress is strongly mediated by intracellular iron (21, rev).
4) Growing evidence proves that Drugs that capture iron (chelators) “kill” cancer cells, without causing damage to healthy tissue, which makes them potentially the most effective antitumor agents (Pahl, 2005).
5) Phlebotomy epidemiologically reduces the risk of acquiring cancer in the general population: Cancer: a Ferrotoxic Pathology (Edgren, 2008; Toyokuni, 2009; ref. 54, 1, rev). In addition, by mobilizing iron –reducing its tissue “sequestration” by the excess of the Hepcidin hormone, it can reduce anemia in critically ill patients https://journals.lww.com/ccmjournal/Abstract/2008/08000/Phlebotomies_or_ery thropoietin_injections_allow.25 .aspx.
6) Chronic anemias do not benefit from the exogenous supply of iron, as long as the INFLAMMATORY / Infectious cause that originates it is not solved, since it is an anemia caused by maldistribution of iron (which does not pass into the blood due to being retained in liver cells and throughout the reticuloendothelial system). Iron- rich diets, by increasing hepatic iron retention, promote greater tumor anemia, due to its Hepcidin-dependent sequestration https://ashpublications.org/blood/article/ 105/4/1797/20350/Hepcidin-excess induces-the- kidnapping-of-iron.
7) Excess iron reserves are a predictor of Hidden Diabetes Mellitus, of Early Coronary Disease, independently and early (262), and of poor prognosis and mortality in subjects with cancer, particularly of the pancreas (263).
8) Epidemiological, experimental and clinical evidence proves it conclusively: processed animal iron causes human cancer (264, 265).
9) The treatment of Chronic Inflammatory Anemia has radically changed; and it is NOT with Iron Supplements, but with Vitamin C, Zinc, which facilitate its mobilization, and adequate redistribution; but, above all, looking for the Causal Disease (266), in its very advanced stages, from Parasitosis, a chronic infection, to Cancer.
10) Regardless of Central Obesity, the accumulation of Visceral Fat directly increases the risk of intestinal cancer (267) and its presence; and this, with causal and irrefutable evidence.
11) Growing accumulative evidence demonstrates it: calorie restriction (without malnutrition) promotes cancer cell apoptosis; and its invasiveness (reducing its angiogenesis) (rev.268, 190) (Fig. 2); likewise, depletion of body iron through blood donation powerfully reduces cardiovascular disease (269), especially the risk of heart attack (156) and, probably, stroke. (A high intake of iron increases the risk of ischemic attack to the brain (270). There is evidence that iron in neonates accelerates brain aging, by causing neuronal death (271); and iron not only free (20) but also accumulated (272) powerfully activates the main intracellular inflammatory and carcinogenic gene: NfkB (20, 27, 272).
12) The greatest risk for acquiring cancer is not genetic: it is the metabolic- hormonal profile (273); and its nutritional overload of iron (1) and other metals (189): accumulated iron directly induces tissue injury (274), and specifically alters the chromosomal activity of oncogenes (275).
13) The fine regulation of iron metabolism (276) and its accumulation (274) is decisive for the extension and prognosis of epithelial cancer.
14) Frequent removal of iron by phlebotomy (1, 277) - blood donation may decrease the risk of malignant tumors in the general population; and it reduces the risk of cancer in “ordinary” humans with insulin resistance (277).
15) Physiopathologically, free iron is harmful, and confers, particularly, greater neuronal (255, 278, 257) and cardiac -cardiovascular- (210, 269) toxicity when it accumulates more in the tissues: and this is much more significant before the Zinc nutritional deficiency (279), a common event in our pregnant population.
16) The progression of a disease, from cancer to a serious infection (eg cerebral malaria) can be substantially alleviated thanks to iron chelation: the capture of the metal (280, 281, 45, 46, 56, 57, 60), its removal (phlebotomy/donation) (277, 1), or an iron-deficient diet (281) have been shown in animals and humans to inhibit tumor proliferation. (as well as therapy-induced cardiotoxicity) (280, 277), and diabetes mellitus (107).
17) How can an optimal energy / protein intake be achieved for each patient, but without intensely stimulating the progression of the cancer? (282): with adequate restriction of inflammatory protein and fat calories, controlling the excessive secretion of INSULIN and its resistance induced by iron (31, 107); the quintessential nutrient that immortalizes cancer cells (283).
18) Free Iron increases the instability of the genome (41, 284) and the chromosome rearrangement, promoting the mutation of proto-oncogenes and the inactivation of cancer suppressor genes (41). Its overload alters DNA and is the cause of specific, powerfully oncogenic mutations (189, 284).
19) Epidemiological data in humans reveal the current evidence, as of 2014: greater exposure to exogenous iron or its overload correlates with an increase in the genesis of cancer (285): thus, metal supplementation, if absolutely necessary in periods critical, should be limited ONLY to extended treatment periods; It can be COUNTERPRODUCTIVE in the long term: iron metabolism must be extremely controlled so that cancer does not spread remotely.
20) We cannot continue "feeding cancer", except when it is incipient; worse if it is advanced: it is scientifically possible TODAY to stop its progress by reducing the abdominal (visceral) accumulation of fat (286) with the reduction of exogenous iron and excess insulin, the generator of de novo cancer (287).
21) The storage of iron as Ferritin in serum, is initially protective and antioxidant; but, in the presence of cancer, when it is excessive, it confers greater aggressiveness to the tumor: High Ferritin is responsible for the progression of cancer, and especially its resistance to treatment (288, 28). It is up to us to begin to abort the greatest epidemic development of cancer, with Comprehensive Oncology Medicine TODAY.
22) Iron is an extremely reactive and oxidizing metal, and its physiological overload potentially increases a greater risk of cancer by directly causing genome instability, so it is essential to modify the current recommendations on its preventive intake, which can become in harmful (289). Cancer treatment –not more important than its prevention- should focus on locating DNA damage in the genome caused by high oxidative stress dependent on iron (290) and copper, metals whose “normal” levels become toxic and promoters of disease (291, 31, 95).
23) At the end of this review, it is verified in Spain, after 59 studies epidemiological in 18 years, that for each mg. Additional animal iron increases the risk of human cancer (292). The higher the iron intake, the higher the insulin resistance and the higher the risk of cancer, and this seems to be more relevant in women (293), as is the regression of breast cancer after intermittent calorie and iron restriction (294).
24) At the end of this review, highlighting the clear and complex interaction between glucose and iron (95), it was shown that glucose loads ONLY TEMPORARILY reduce serum iron concentration (Aigner, 2013); (causing confusion about their levels); and this by increasing its tissue sequestration due to an elevation of hepcidin, the iron regulating hormone (295).
25) Iron loads promote profound deleterious changes in the proteins (cyclins) that control the cell cycle (296, 58, 59), potentially generating cell malignancy. And among all the metals in contact with man, iron is the one that most permanently modifies the genetic material, its unbalanced contribution (due to the lack of natural Zinc, for example) being the initial cause of carcinogenesis. and aging (297, 189).
26) Liver cancer (Hepatocellular carcinoma), the third leading cause of cancer mortality, especially among developing countries, can be clinically reduced with phlebotomy, since iron overload is a significant factor for its development: and deprivation iron is proving to suppress cancer growth (Ba, 2011) (298, 285, 101): free iron can be cytotoxic and genotoxic, in the central nervous system (299).
27) The experimental evidence that began 30 years ago is categorical, although unfortunately neglected by “great researchers”: cellular iron deprivation powerfully reduces the proliferation of tumor cells (280, 301); which initiated the most effective anti-cancer treatment: iron depletion and chelation (45-47, 53, 56, 60-63); which also prevents the usual cardiotoxicity of chemotherapy, as well as super-infections that are almost always fatal (280).
30) There should not be an Iron/Zinc imbalance in favor of nutritional iron; given their complementary and antagonistic physiological actions, especially in the face of carcinogenesis: any deficiency, even zinc deficiency, will increase tissue accumulation of iron (306) and its cytotoxic, inflammatory and carcinogenic potential: its chronic accumulation definitely causes organic damage (pantropic) subclinical and unappreciated by the current medical community (307, rev).
31) Genetic inheritance (Genotype) does NOT determine the appearance of Cancer: it is the conjunction of Metabolism and Environment (Nutritional and Physical) that finally determines the PHENOTYPE that will signal the appearance and aggressiveness of cancer: This is the forgotten basis of the interaction Nutrients-Genes. When the production of glucose exceeds its use (insulin resistance) the permissible biological system for the appearance of cancer is constituted (308, rev).
32) Yes, as we have seen, excess iron perpetuates the immortality of the cancer cell (powerfully inhibiting its programmed cell death directly or through the resistance (excess) of Insulin, in cells for specialized normal cells, excess iron (like calcium) induces their degeneration and death (like neuronal degeneration) (309; 257, 278, 261).
33) At the end of this work, it is verified, in a very well achieved meta-analysis (449 articles and 11 prospective studies) (up to the year 2012) that the high consumption of Heme Iron and/or the increase in the body iron reserves are significantly associated with a higher risk of suffering from Diabetes II (310, rev); which confirms, once again, the Diabetes-Cancer relationship (and vice versa).
34) The greater the "sequestration" of iron in the cell, the greater the severity of a cancer: the lower the amount of iron exported from the intracellular (the hormone hepcidin inhibits its cellular efflux by degrading ferroportin), the greater the aggressiveness of the cancer in women (as confirmed in incredible genetic studies for breast cancer) (311, ed).
35) In diabetes and hematopoietic cancer in particular (leukemias) (and in any neoplasia and cancer in general) free or isolated iron should not be given because it increases microbial growth and profoundly damages phagocytosis (altering neutrophilic lactoferrin, responsible for capturing iron and reducing its bioavailability for the bacteria) (312, 313).
36) An excess of iron, especially in the face of any biological stress, affects the physiology and leads to cell injury (314, 315). Given the direct evidence that its regulated deprivation induces apoptosis in animal lymphoma, it constitutes a safe and effective rational strategy for the treatment of human cancer (Kovar, 1997) (316, 1); and especially when it has been shown that the higher the dietary intake of iron, the greater the risk of invasiveness of human cancer (317). And, like unbound iron, free (or excessive) hemoglobin is highly oxidizing and toxic to cells and tissues, so that its reduction by means of a restricted supply of iron decreases the size of the tumor, especially if it is malignant (318, 319, 320).
37) After the publication of this review article, it is verified once more now in a very large Asian cohort of more than 300 thousand adults 81997-2008), and in populations without a family history of cancer, that elevated serum iron is today a common disorder and a risk marker for cancer (321, 322). Furthermore, even among the population with cancer, blood donation significantly reduces mortality, which has been solidly proven in 20 years of epidemiological studies (“healthy donor effect”): better health with blood donation will be achieved even among cancer patients (323); it is evidenced that, in particular, reducing the excess of neutrophils will reduce the growth of metastases and their “novo” generation https:// www.frontiersin.org/articles /10.3389/fimmu.2020.565165/full
38) Solid and growing epidemiological, experimental and clinical evidence prove to this day that: only a discrete increase in exogenous iron contributions increases the growth of any tumor, significantly raising the risk of cancer occurrence, and especially its mortality (23, 324, 325).
39) Without fear of being wrong, we can affirm that, in both men and women, chronic iron excess is even more mutagenic and oncogenic than cigarette smoke, being powerfully synergistic in accelerating cancer disease: iron it is demonstrably the major regulator and promoter of the cell cycle (244; 322, 326).
40) Reductions in blood iron concentrations prevent cancer morbidity and mortality (327, 328), since the higher the free iron concentration, the higher the incidence of cancer in humans (165, 317).
41) Iron facilitates the evasion of the tumor to its eradication by the immune system: that is: the reactive metal protects the cancer cell from its immunological destruction (329). Therefore, no cancer can be effectively eradicated if high levels of tissue iron persist.
42) Iron supplementation is currently ineffective, and potentially harmful to health; being profoundly so in the presence of obesity or sedentary lifestyle (330). Let us remember that the permanent recycling of 95% of body iron (Heme iron from animal sources) is the most powerful energy source for the development of the cancer cell and for any neoplastic disease, constituting its greatest risk for its prevalence, severity and mortality (331).
43) Conclusively, it is confirmed that the higher the accumulation of body iron or free circulating iron, the greater the risk of acquiring diabetes, and of dying from cancer (332; 333); and the greater the exogenous supply of vitamin C (its natural chelator), the greater the protection against genotoxic and mutagenic damage from free iron (334; 317, 159, 283,165, 1).
44) A patient with cancerous or hematological disease will have a worse prognosis the greater the accumulated iron present; and transfusions can be extremely deleterious, as they cause potentially fatal complications: the major malignant transformation induced by iron, proven epidemiologically (335, rev) is reduced with the decrease of the metal (1).
45) Particularly, the dietary excess of iron and arsenic in a context of Deficiency of anticancer micronutrients (Omega-3, Zinc, Selenium) is one of the most powerful factors in the promotion and extension of cancer pathology due to causing genome instability, irreversible promoter of malignancy (336). Convincingly, the contributions of iron (and its metabolism) are the final determinants in the synthesis and repair of DNA (336, 337, 41, 284, 1, 240); and today, crucial not only in the promotion but also in the initiation of cancer (338, 339). (An increased risk for acute myeloid leukemia in Down syndrome has even been reported in relation to iron supplementation and multivitamins, Blair, 2008) (340). This shows once again that excessive iron is crucial for the development of cancer (36.1). And this is solidly proven in hepatocellular cancer (341, 342), where iron-enriched dairy products potentially increase tumor size; on the contrary, its restriction reduces the advance of the cancerous process, by reducing the proliferation of malignant cells (343, 344).
46) Specifically, the contribution of iron suppresses the programmed death of the tumor cell (344, 1, 47) and its restriction limits the growth of the tumor mass (345, 47, 1) (American Society of Hematology, 1990) and the extension of the cancerous process. Let us not forget this overwhelming evidence; even so, iron must not accumulate and instead must be “exported” out of the cell, otherwise it will cause malignancy (346, 347, 348, 280, 278, 1).
47) Precisely, the ferritin measured both in the serum and tissues, is it constitutes the most effective predictor for the diagnosis and prognosis of cancer (349, 350); since tissue ferritin directly stimulates tumorigenesis (351, 287): The integrity of the genome is established and determined by our diet (288, 352, rev). And if man donates blood, he will reduce his increased risk of cancer (353, 354, 10, rev), as demonstrated by recent rigorous epidemiological studies in humans and animals (354, rev; 355, rev), while reducing oxidative damage to your DNA, even in optimal health (356).
48) Chemotherapy and radiotherapy, which have been shown not to eradicate the cancer, are and will be ineffective as long as cancer-initiating cells/Stem Cells are maintained, which will survive as long as high levels of cellular iron are maintained; Thus, it has been confirmed in humans that iron promotes greater cancer aggressiveness by inducing Stem Cells, that is, the root of cancer (357), and its accumulation powerfully induces greater mortality (335, 10, 1, rev).
49) Dietary isolated, and molecularly free (catalytic) iron is a generating or inducing potential for spontaneous mutagenesis, whether initiating or promoting cancer (358, rev): and the lower its acute exogenous contribution, the lower the inflammation in the microtumoral environment, clearly promoting cancer (20, 16, 359, 1), where iron not only contributes to the progression, but also to the initiation of cancer (337, rev, 360, 1, 44, 56, 361, 362, rev), especially hematopoietic cancers - such as leukemia and myelomas - (363).
50) Intracellular capture of iron not only alters the cancer cell, but affects its inflammatory microenvironment: If we want to fight cancer, we must modify the histologically proven “cancer” biology of iron (364, 365), given its powerful role as the metal in stimulating and maintaining the most common cancer signals (366, 1, rev); Otherwise, any chemotherapy or radiotherapy will increase the resistance of the tumor, by increasing the inflammatory signals that feed the cancer (367, 66, 287, 368, 362, rev, 202); and this because it powerfully increases the bioavailability of free iron (369, rev). Loaded iron potentially induces seizures due to its high neuroinflammatory power and should be outlawed in epilepsy and in the entire field of neuro-oncology (370, 371).
51) The progression of the Cell Cycle that occurs in normal tissues, and particularly during the development of tumorigenesis is finely controlled by anti-apoptotic signaling from iron, an essential nutrient for the aberrant and uncontrollable cell proliferation (372, rev) that occurs in cancer.
52) An optimal immune function is initiated and determined by an optimal cell metabolism (373, rev); continuous or excessive iron intake alters it negatively, reducing physiological immune-protection against cancer (328, 160, rev; 63), in addition to altering the Insulin axis, the master hormone that regulates anti-immunity (374). -neoplastic.
53) The energy that powers cancer cells (glycolysis) increases in the presence of inactivation of the anticancer gene p53 (375), which is strongly increased by dietary iron overload: thus, heme iron binds, interferes with, and degrades the major tumor suppressor protein P53 (376, 377, rev); while its deprivation, by stabilizing it, suppresses the dependent tumor formation, thus constituting the best coadjuvant treatment of the isolated and obsolete current chemotherapy (377 rev). This demonstrates that free iron (or its dietary overload) is a promoter of pre-neoplastic fibrogenesis (378, 379, 1, 10, 380, 356, rev), powerfully promotes cancer cell survival (381), and the molecular mechanisms that initiate, promote, and sustain cancer (385, 386, 382, 1, rev; 383, rev, 384, rev), as oxidative damage to DNA -8-OHdG- is controlled by body iron stores (386).
54) Blood extraction decreases the incidence of visceral cancer, as well as its mortality, in humans (387). Repeated phlebotomies are being shown in vivo to reduce the size of the cancerous tumor, its histology and malignancy, as well as the magnitude of its extension (asbestos-induced mesothelioma) (ref. 388, figure 13). If cancer cells require higher amounts of iron than normal cells, phlebotomy will be the best adjunct not only to prevent but also to reduce the extent of cancer disease (138; 388, rev), by improving iron redistribution and iron circle vicious that perpetuates systemic inflammation (389, rev) that perpetuates cancer, and accelerates the presentation of its main risk factor, diabetes mellitus (187, 184, rev).
55) Iron (89, 101, 105, 197, 114) oral or intravenous pharmacological (390) promotes death and apoptosis of beta cells and causes diabetes or accelerates and aggravates its complications (105, rev), since the potent mutagenic effect of excess free iron only occurs in the presence of stable and normal oxygen concentrations (391), unlike its unique necro-apoptotic cytotoxic effect, in the face of hypoxia-hyperoxia patterns (256, 392-398).
56) Due to all that has been extensively documented, serial phlebotomies -better from adolescence- will be a magnificent therapeutic weapon (alone or together with iron chelation) to avoid the complications and severity of chronic metabolic and oculo-neurodegenerative diseases (399), from diabetes (31, 94, 106, rev) to cancer (1, 10, 39, rev).
57) All chronic Anemia is of Inflammatory cause, and is fundamentally due to excess Hepcidin, which "kidnaps" Fe in tissues. (1, rev, 400, rev,401, rev, 402): therefore, anemia is not due to a lack of iron, but to its deficient distribution promoted by inflammation and/ or hidden infection. The doctor runs a high risk of worsening the disease, by administering iron: eg, the greater the intracellular iron deposits, the greater the serum ferritin -not nuclear, which protects DNA-, the greater the resistance of the cancer cell to its total eradication (403, 404). It is incredible that, until five years ago, the numerous physiological damages that free iron causes by itself and in combination (401), (Kell, 2009: extensive rev. 2469 references) have not been appreciated. Dietary iron and insulin resistance (occult hyperinsulinemia) together constitute the greatest risk for cancer, which, however, can be prevented with frequent blood donation or phlebotomy (405, rev). In cancer, exogenous iron will promote greater tumor anemia (69, rev).
58) It is essential to correct the altered metabolism of cancer cells that initiate cancer (Cancer Stem-Cells), even before the genomic alterations (406, 1, rev) -promoted by the inflammatory microenvironment, originated and sustained by excess of iron or glucose - to eradicate the disease. That is to say: If we do not control the metabolic alteration that precedes (406, 1, rev), sustains and feeds the permanence of the Mother cells (Cancer Stem-Cells) at all stages of the carcinogenic process , the cancerous disease will never be eradicated (406 , rev, 407, rev, 408, rev, 409), and it will always recur, since it is cellular iron that allows the survival of these cancer root cells and all neoplastic cells (409, 24, 25, rev)
59) We summarize overwhelming evidence: exogenous iron will only worsen the chronic disease -chronic inflammatory anemia- by promoting greater hormonal sequestration of iron (410, 411, 168), and aggravating the disease (331, 332, 324, 309, 313, 314, 323, 326, 335, 337, 290, 360, 372, 380, 385, 339, 356, 316, 317, 318, 284, 237, 251, 255, 259, 264, 268, 269, 292, 291, 276, 279, 399, rev, 8, 9, 26, 29, 40, 69, 71, 83, 89, 92, 121, 412, 1, rev; 413) and its high risk of infection (414, rev) and diabetes. In the face of any childhood anemia, an infectious process should be sought; and after treating it, the anemia will be spontaneously corrected in less than two weeks (Franco-Tamayo, Anemia in the Colombian Infant Population-Anemia Working Group Latin America- Rev, AWGLA, 2005: Vol, N2).
60) Today there is no doubt that iron deposits accelerate diabetes; and vice versa: phlebotomy reduces pancreatic fibrosis, Glycosylated Hemoglobin, raising basal insulin secretion (415), improving its inflammatory-metabolic-vascular complications. And if you don't have a history of diabetes, iron loading may initially “improve” your glucose tolerance (415), by reducing its hepatic output (416), but will speed up your molecular pathway (Akt Signaling) to liver damage., hidden diabetes and cancer (416).
61) Iron initiates cancer (337, 417, 361, rev), spreads it (1, 337, rev) and promotes their resistance to treatment by inducing the genes and organization of Cancer Stem Cells, and stimulates their high degree of malignancy, and their devastating aggressiveness (417, 357).
62) Iron overload or daily iron in cancer patients independently increases tumor growth (418, 419, 420, 360, 362, 376, 377, 1, 2, 3, 4, 6, 8, 25, 421, rev, and 70 ref.) and has directly demonstrated its carcinogenic initiating power, by causing dysfunction of the main cellular epithelial adhesive (stabilizing) protein (e-cadherin) (422, 423), and by stimulating the major family of oncogenic proteins Myc (424, 425, rev). And on the contrary: the capture (binding) of iron with great affinity, by the apoptotic glycoprotein Lactoferrin, inhibits (in vitro, in experimental and clinical studies) the extension and aggressiveness of human cancer (321, 337, 426, 427, 428, 429, 430) and its high metastatic power (428,429).
63) If the apoptotic, anti-carcinogenic, and anti-inflammatory power of Lactoferrin is inhibited and becomes mitogenic and proliferative, when it is saturated with Iron (428, 431, 432), it is demonstrated, once again, the inflammatory power of intracellular iron, which increases susceptibility to infections and concomitant chronic anemia (433, rev, 411, rev). Along with quarterly phlebotomy, human lactoferrin in maternal colostrum will reduce cancer. The molecular evidence - in vivo- and in vitro, is conclusive (434; 435).
64) Exogenous iron in supplements is Inflammatory (433, rev) (Intestinal immediate and cumulative (436, 437, 438, 439, 440), genotoxic and pro- tumorigenic in vivo (159, 334, 440) due to its gene suppressive power most important anticancer -p53- (376, rev, 377, rev) and its high capacity to stimulate metastasis (441, 442) Transfusion loads can promote clonal evolution towards acute myeloid leukemia (445, rev)
65) Every disease, and especially cancer, appears and remains within a profound alteration of cellular metabolism (443, rev), sometimes irreversibly altered by exogenous iron, a proven inflammatory toxin, initiator, promoter, accelerator of the cell cycle (446, rev, 447, rev) and generator of malignancy (448, rev, 449, rev), altering the protective activity of the main cancer suppressor gene, and which provides the greatest genetic barrier for neoplastic transformation (450 , 451, rev). While this protective gene is the guardian of the genome, its mutation is the guardian of cancer (1, rev, 337, rev; 451, rev).
66) Established the inhibitory relationship P53-IGF-1 (452, rev), if the p53 gene inhibits the proliferative and carcinogenic Axis of Insulin-IGF-1, and decreases glycolysis; and cellular iron inhibits the p53 gene or causes its mutation, an increase in catalytic iron turnover powerfully activates the survival and induced nutrition of the neoplastic cell (the example of the increase in lung cancer by iron and the vitamin retinol derivative A preformed (457, 458).
67) The higher the ingested iron, especially in the presence of our common inflammatory diet, the greater the risk of the appearance and aggressiveness of any cancer (458, 459, rev, 1, rev, 74), given its evident ability to start the cancerous disease early (460, rev) in addition to its powerful promoting action, especially when faced with high amounts of oleic, palmitic (461, 462; 463) or omega-6 (464, rev, 465, rev) acids; with the exception of the omega-3 anti-inflammatory fats, with proven molecular (464, rev, 465, rev; 466,467) and clinical (468, 469) anti-carcinogenic effects, as well as zinc, a trace element of greater nutritional importance to maintain genome stability (305, rev, 470, rev, 471, 472, rev, 237, 278).
68) Overexpression of cellular Ferritin increases expression of the oncogenic/ promitotic factor FoxM1, an inducer of the epithelial/ mesenchymal transition (480, rev). Therapeutic phlebotomies reduce the risk,and are proving to reduce the spread of cancer, especially liver cancer (484, rev; 487). Iron excess is evidently the greatest risk in the generation of cancer and in the causation of preneoplastic lesions; its mobilization or capture together with the restriction of caloric energy is essential for its eradication (489, rev, 490). The evidence is growing, but still ignored by hidden and petty interests (476, rev, 491, rev, 501, rev). –This is corroborated by new evidence: the polyphenol Curcumin reduces the spread of cancer by inhibiting the cellular accumulation of iron and chelating the metal (494, 495, rev), being able to reduce the aggressiveness/ extension of leukemias, clearly increased by the genomic instability promoted by iron overload (502, rev): The higher the serum ferritin, the greater the intensity of childhood cancer aggressiveness (503, 81, 82,139). While the incidence of cancer increases rapidly and markedly in recipients of a blood donation, among those donors, the incidence significantly decreases (504, 505; 42). If macrophages and neutrophils promote cancer development; and conversely, neutrophil depletion inhibits lung metastasis in vitro, iron donation, by reducing its inflammatory plasticity, will reduce carcinogenesis (Liang, 2021) (509, rev).
69) Excess iron cannot be excreted or eliminated: the forgotten pathophysiology (Jung, 2019, rev) (511, rev)
"Iron must be transported, balanced, used and eliminated, but never accumulated, neither in our daily life, nor in our transitory death"“To rapidly improve symptomatic chronic anemia, administer only Vitamin C will immediately mobilize your sequestered iron” Jara J, 2019; Toyokuni, 2017
https://www.sciencedirect.com/science/article/pii/S0891584918317180?via%3D ihub#bib163 Badu-Boateng, 2019, rev
Figures
Figure 1: Meaning of the Powerful Oxidation Effect induced by Iron in the Genesis of Cancer.Schematic View (Taken from: Tokoyuni S, 2009) (1).
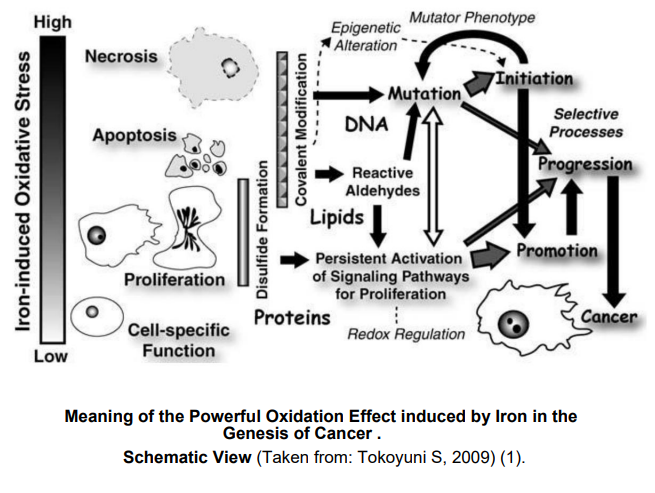
Figure 2:Calorie Restriction and Cancer:
A) Caloric Restriction (CR) effectively inhibits several types of cancer in animal models.
B) Correlation between the lowest incidence of Tumor and the degree of Restriction caloric.
C) Demonstration of parallel and opposite effects of CR on cancer and aging: the ability of CR to decrease the Insulin-IGF-1 axis (strongly potentiated by Iron), and the inhibition of PI3K-AKT insulin signaling, simultaneously protect cells from aging and cancer. (Taken and modified from: Pallavi S; 2012) (268).
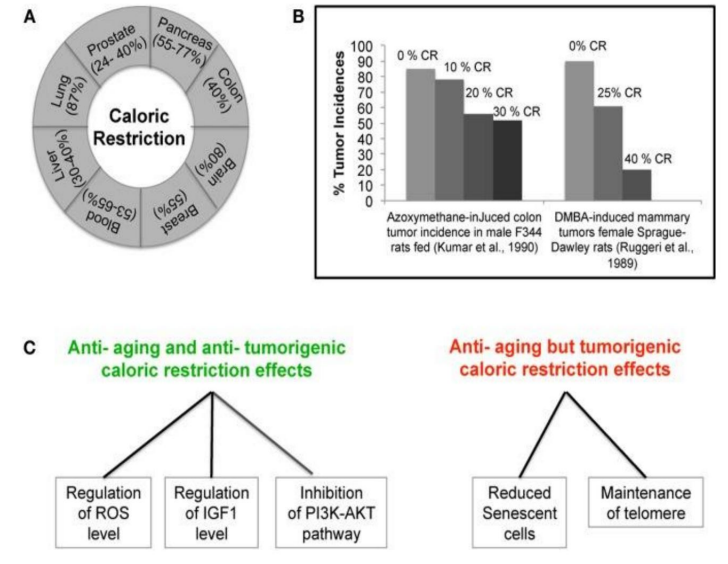
Figure 3: Simplified scheme of the pathways by which Iron Induces Diabetes (Taken from Swaminathan S, 2007) (106).
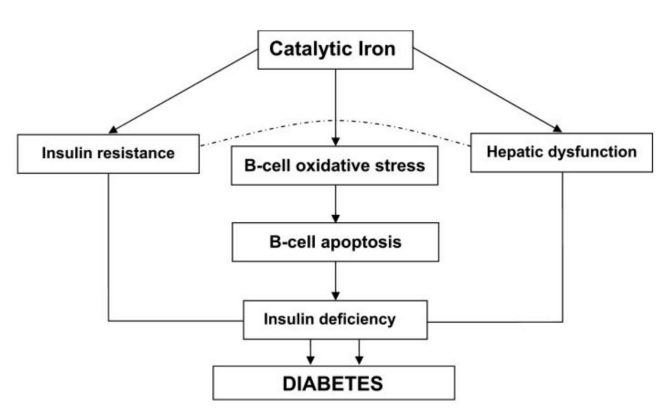
Figure 4: Ferritin concentrations in breast aspirate (areola). Substantive differences between the mammary fluids of women without or with cancer (Taken from: Mannello F, 2012) (273)
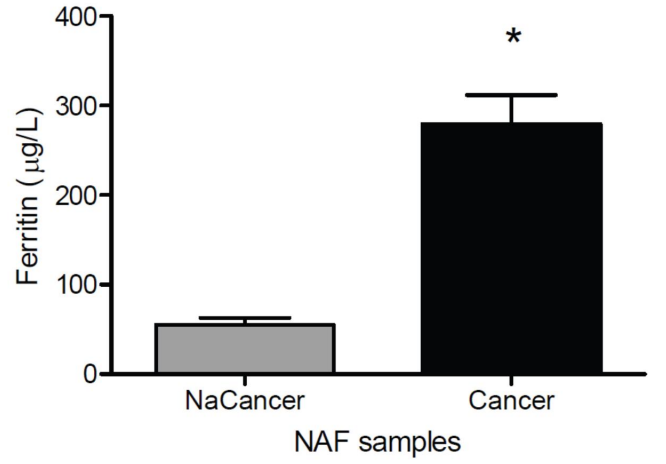
Figure 5:Ferritin Concentrations in Breast Areolar Aspirate Fluids In Women with or without cancer, according to their menopausal status (pre or post-menopausal). (Taken from: Mannello F, 2012) (273)
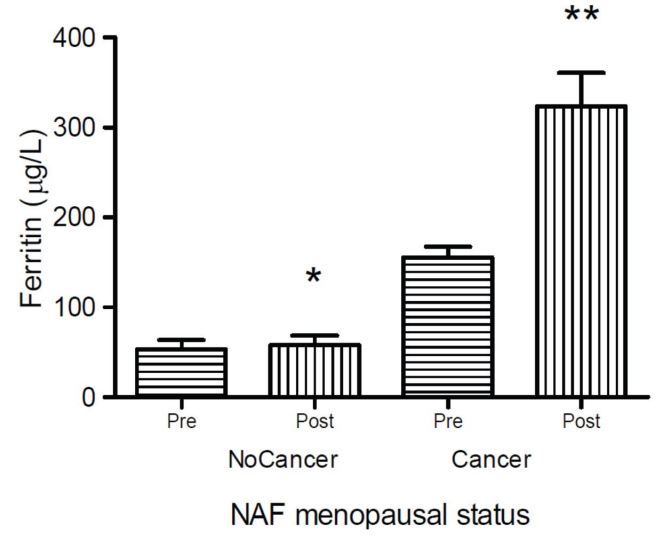
Figure 6: Scheme that demonstrates the actions of the Hepcidin hormone on the metabolism (sequestration) of Iron in the tissues: The Increase of Hepcidin - before any inflammation / infection or advanced cancer - is the fundamental cause of Chronic Anemia, therefore resistant to the contribution of Iron. Modified from: Ganz, 2012 (168)
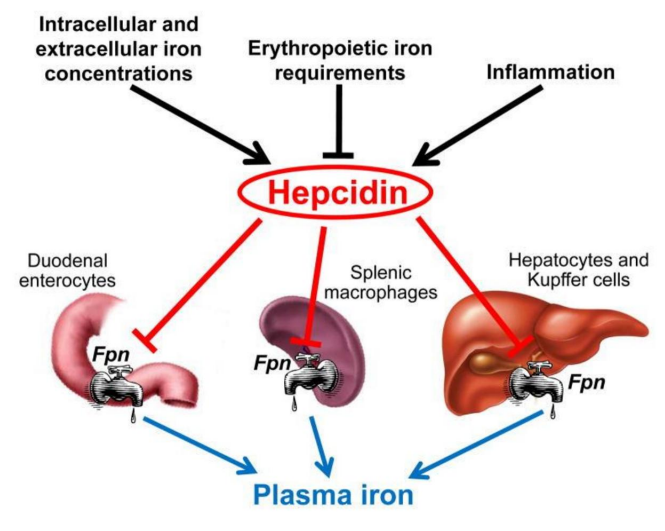
Figure 7A: How mammary tumor size progressively increases in rodents with the addition of lucose, insulin, or both. Figure taken, with modified text from: Vigneri, 2009 (176, rev)
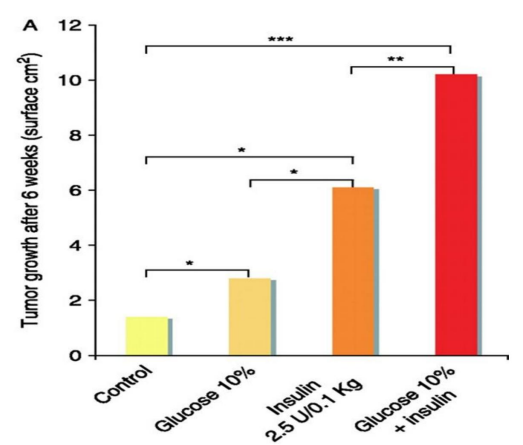
Figure 7B: Reduction of tumor in acute chemical reduction of insulin.It is well established that: tumor size ill increase with the addition of oral iron (not shown in chart-A)Figure taken, with modified text from: Vigneri, 2009 ( 176, rev)
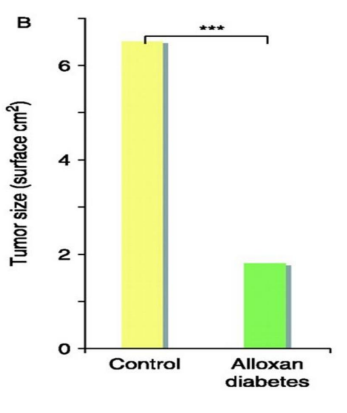
Figure 8: Diagram showing the different mechanisms by which iron promotes oxidative stress that accelerates aging diseases in particular; generating an accumulation of non-degraded toxic products (denatured proteins) and lipid peroxidation; and its direct damage to cellular and mitochondrial DNA, all of which could be stopped (left side of the figure) in the event of a deprivation of free or catalytic -labile- Iron, which will be achieved through a regular extraction of iron (by donation , chelation or restriction). Otherwise, excessive oxidation to proteins (leading to their alteration and accumulation) will override any natural DNA repair mechanism.Modified from: Galaris, 2008 (300, rev)
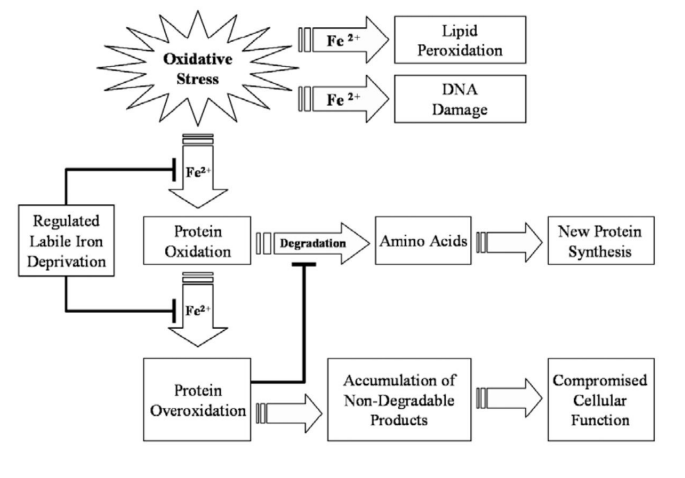
Figure 9: Image showing the Activated Signaling Patterns in the various types of Cancer caused by excess Iron We will ention only two molecular signals that initiate oncogenesis: Cell Cycle Progression that does not kill malignant cells, and the inhibition of the P53 gene that activates apoptosis (and “kills” cancer cells). (For further details, see Text) Taken from: Zhang, 2015 (321, rev).
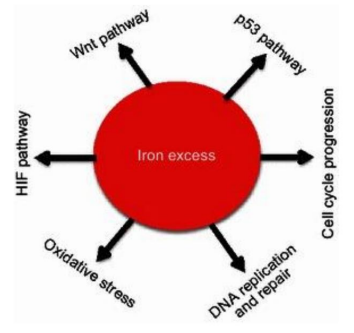
Figure 10: Molecular schematic showing how cellular iron determines metabolism and genome and chromosome integrity (explanation in the text: Torti, 2013) (337, rev).
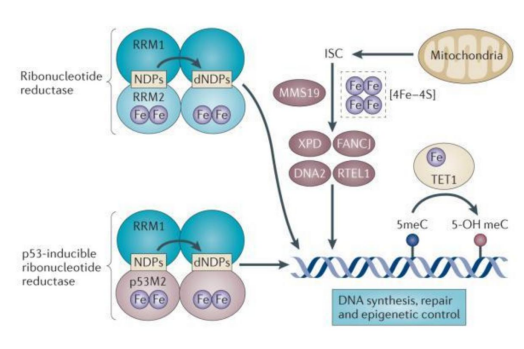
Graphic 1: In the general population, all-cause mortality is shown to increase significantly with higher transferrin saturations (panel B): and it is the greater increase in non-transferrin-bound toxic iron (transferrin saturation >60%) that is the factor that increases said radius in mortality (Taken from: Puliyel, 2015) (ref. 335)
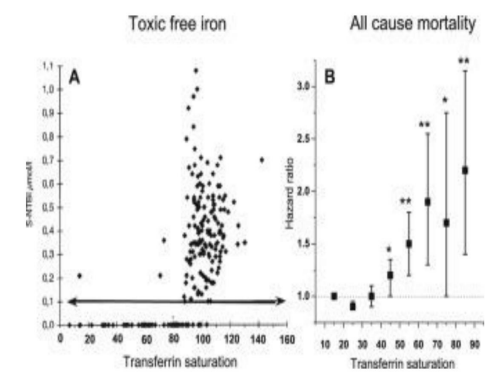
Figure 11: Scheme that shows how Oncogenesis and Cancer are generated through intense mutagenic oxidative damage that promotes genomic instability,and all this is generated by iron overloads, which, through the Fenton reaction, convert Iron to its Ferrous state ( Fe++) to its Ferric state (Fe+++).(Scheme taken with modified text from: Bystrom, 2015) (ref. 362, rev).
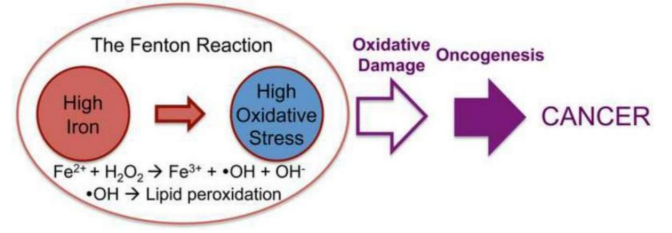
Figure 12: Iron involved in the genesis, progression or maintenance of cancer. Taken from: Zhang, 2015 (321, rev)
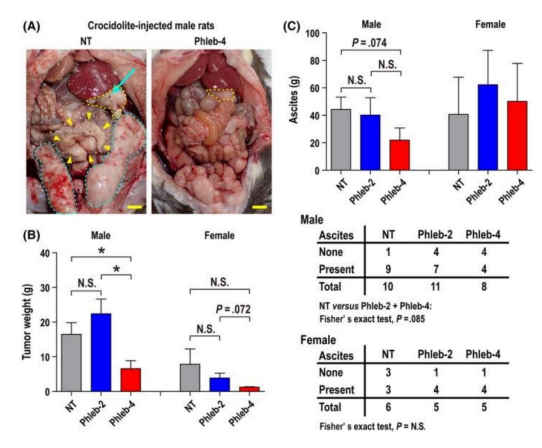
Figure 13: The reduction of peritoneal tumor dissemination of asbestos-induced Mesothelioma in rats is shown: disappearance of metastases in rats with 4 phlebotomies, compared to the control group ( Phleb-4 vs NT).(Taken from: Ohara, 2018) (ref. 388)
In 2020, there is evidence of an increase in the aggressiveness of cancer in Africa due to excess of Iron, which can be reversed with phlebotomies and natural chelators)
Orisakwe OE, Amadi CN, Frazzoli C Management of Iron Overload in Resource Poor Nations: A Systematic Review of Phlebotomy and Natural Chelators J Toxicol.2020 Jan 27;2020:4084538.
Figure 14: Scheme that shows that it is the metabolic decontrol of the root cancer cells (Stem-Cell) that precedes their proliferation, dissemination and resistance to treatment (insulin-mediated anti-apoptosis). (Taken and Modified from: Chae, 2018) (Ref. 408).
Figure 15: It is shown that the main Tumor Suppression Protein p53 is down-regulated by an excess of iron Compared with mice that received a normal (normal-low) iron diet, those that received a high-iron (double) diet had significantly reduced levels of the tumor suppressor protein p53 (Figures 1D and 1E.It is concluded that diets high in iron (resulting tissue overloads) are causally correlated with a significant decrease / inactivation of the anti-tumor protein p53,which directly stimulates the invasiveness - mesenchymal-epithelial transition - of Cancer (Ying, 2021, rev) (ref. 510)(Bhutia Y. Chronic exposure to excess iron promotes EMT and cancer via p53 loss in pancreatic cancer. Asian J. Pharm. Sci. 2020 (ref. 513). Taken (with modified text) from: Shen, 2014 (detailed explanations: Ref.276).

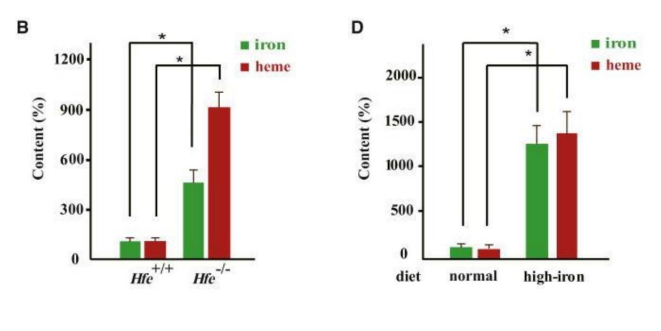
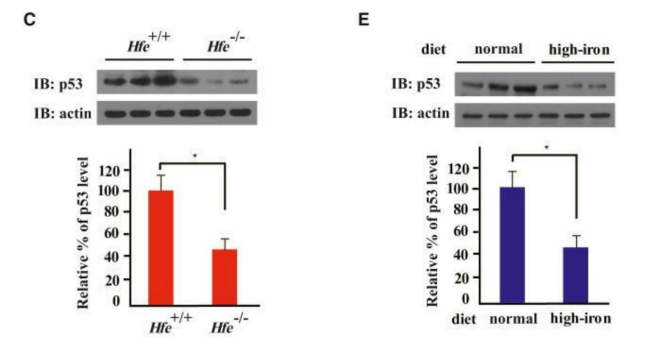
Figure 16: Histological images of the distal colon that demonstrate the inflammatory effect of oral iron (in the photos,in an induced colitis)
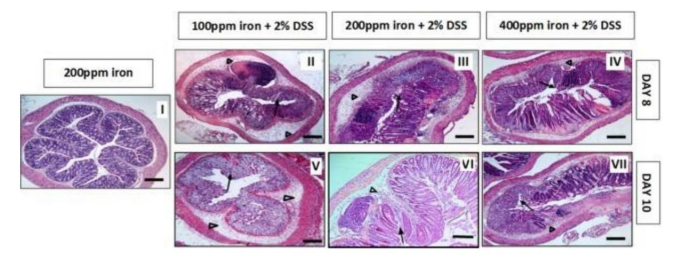
Image and text taken from: Mahalhal, 2018 (ref. 439)Taken (with modified text) from: Shen, 2014 (detailed explanations: Ref. 276).It can be seen how the submucosal edema increases as iron increases (arrows) and the almost complete loss of colonic epithelium, as the exposure time to the high iron diet increases.Mice received Water (control group I) or Dextran Sodium Sulfate for 5 days, followed by: another 3 days of water (see how dissolved iron increases) (boxes II, III, IV), or 5 more days of Water (V, VI,VII)It can be seen how the submucosal edema increases as iron increases (arrows) and the almost complete loss of colonic epithelium, as the exposure time to the high iron diet increases.
Image and text taken from: Mahalhal, 2018 (ref. 439)
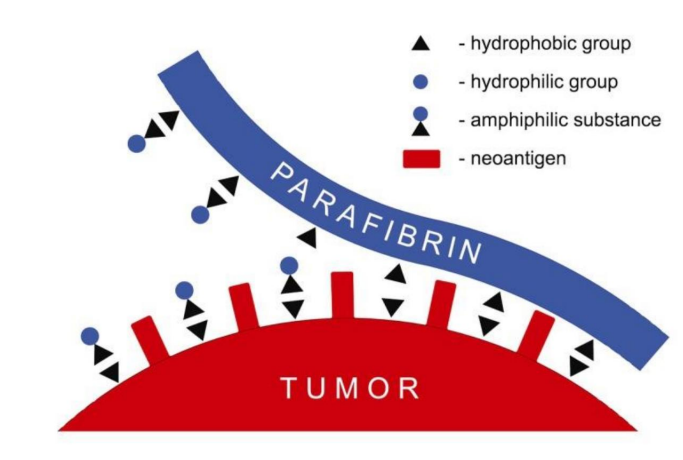
Figure 17: It is shown how the Parafibrin formed by free Iron (hydrophobic altered plasmatic fibrinogen), totally resistant to proteolytic degradation, powerfully protects the tumor cell from its recognition and destruction by the immune system; and induces a permanent state of inflammation, extension and progression of the tumor Taken from: Lipinsky, 2014 (ref. 446, rev) (further explanation in texts of: ref. 446 and 447, rev)
It shows, graphically, how the extraction of blood (Phlebotomy or Donation) in a Physiological way stimulates Erythropoiesis -and relieves chronic anemia, especially any inflammation and cancer- by suppressing the hormone Hepcidin, which increases bioavailability of iron (by reducing its hepatic sequestration) for increased blood formation.By bone marrow erythroblasts: today duodenal iron absorption increases with phlebotomies Ginzburg, Hepcidin-ferroportin axis in health and disease, 2019 https://www .ncbi.nlm.nih.gov/ pmc/ articles/ PMC7730607/
We demonstrate here that by naturally stimulating the synthesis of Erythrophenone,the regulating hormone of erythropoiesis, the Anemia of Inflammation is largely reversed (453, rev, 454, rev. 455, rev, 456. rev)Graph taken and modified from: Kautz, 2014 (ref. 454, rev).
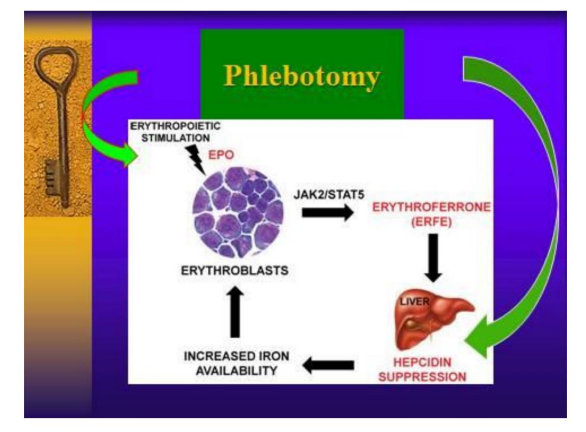
The chart may also explain, in part, why regular blood draws are being shown to reduce cancer prevalence and mortality, as well as being an effective therapy (along with dietary restriction) to arrest and reduce the disease (455 , rev, 387, rev, 447, rev)Excessive iron affects bone marrow and damages hematopoiesis (473, rev, 474, 475).Precise sequestration of intracellular iron (within lysosomes) in conjunction with caloric restriction destroys Cancer Stem Cells (Mai, 2017; Mihaylova, 2014)(ref. 479, 488, rev)
Today it is strongly evidenced that iron is essential for the initiation and survival of cancer-initiating cells -Cancer stem cells-. And this explains its earlier appearance in each generation.
(Torti, 2020) (ref. 496, rev; Ogino, Nature Reviews Clinical Oncology rev). The epidemic of early-onset cancer https://mail.google.com/mail/u/0/?ogbl#inbox/FMfcgzGqQckHhPxbWDbRGFPkhBSk
References
- Toyokuni S (1996) Iron-Induced Carcinogenesis: The Role of Redox Regulation Free. Radic Biol Med 20: 553-566.
- Awai M (1989) Pathogenesis and mechanism of iron overload: ferric nitrilotriacetate, hemosiderin, active oxygen, and carcinogenesis Rinsho Ketsueki. 30: 1115-1127.
- Rezazadeh H, Athar M. (1997) Evidence that iron-overload promotes 7,12- dimethylbenz(a)anthracene- induced skin tumorigenesis in mice Redox. Rep.3: 303-309.
- Blanc JF, De Ledinghen V, Trimoulet P, Le Bail B, Bernard PH, et al. (1999) Premalignant lesions and hepatocellular carcinoma in a non-cirrhotic alcoholic patient with iron overload and normal transferrin saturation. J Hepatol. 30: 325-329.
- Okada S (1996) Iron-induced tissue damage and cancer: the role of reactive oxygen species free radicals. Pathol Int 46: 311-32.
- Nahon P, Ganne-Carrié N, Trinchet JC, Beaugrand M et al. (2010) Hepatic iron overload and risk of hepatocellular carcinoma in cirrhosis. Gastroenterol Clin Biol 34: 1-7.
- Bhasin G, Kauser H, Athar M (2004) Low iron diet retards 12-O-tetradecanoyl phorbol-13-acetate-mediated tumor promotion in murine skin. Arch Toxicol 78: 283-289.
- Heath AL, (2003) Fairweather-Tait SJ Health implications of iron overload: the role of diet and genotype. Nutr Rev 61: 45-62.
- Stál P, Hultcrantz R, Möller L, Eriksson LC et al. (1995) The effects of dietary iron on initiation and promotion in chemical hepatocarcinogenesis. Hepatology 21: 521-528.
- Toyokuni S (2009) Role of iron in carcinogenesis: cancer as a ferrotoxic disease. Cancer Sci 100: 9-16.
- ROBINSON CE, BELL DN, STURDY JH (1960) Possible association of malignant neoplasm with iron-dextran injection. A case report Br Med J 2: 648-650.
- CROWLEY JD, STILL WJ Metastatic carcinoma at the site of injection of iron dextran complex.
- MacKinnon AE, Bancewicz J (1973) Sarcoma after injection of intramuscular iron. Br Med J 2: 277-279
- Greenberg G (1976) Sarcoma After Intramuscular Iron Injection. Br Med J 1: 1508-1509.
- Weinbren K, Salm R, Greenberg G (1978) Intramuscular injections of iron compounds and oncogenesis in man. Br Med J 1: 683-685.
- Lund EK, Wharf SG, Fairweather-Tait SJ, Johnson IT et al. (1999) Oral ferrous sulfate supplements increase the free radical generating capacity of feces fromhealthy volunteers. Am J Clin Nutr 69: 250-255.
- Day SM, Duquaine D, Mundada LV, Menon RG, Khan BV et al. (2003) Chronic iron administration increases vascular oxidative stress and accelerates arterial thrombosis. Circulation 107: 2601-2606.
- Polette A, Blache D. (1992) Effect of vitamin E on acute iron load-potentiated aggregation, secretion, calcium uptake and thromboxane biosynthesis in rat platelets. Atherosclerosis 96: 171–179.
- Praticó D, Pasin M, Barrin OP (1999) Iron-Dependent Human Platelet Activation and Hydroxyradical Formation: Involvement of Protein Kinase C Circulation 99: 3118-3124.
- She H, Xiong S, Lin M, Zandi E, Giulivi C, et al. (1997) Tsukamoto S Iron Activates NFkB in Kupffer Cells AJP Gast Liver Physiol 2002 Sep; 283(3): G719-G726 21. Meneghini R Iron Homeostasis, Oxidative
- Stress and DNA Damage. Fr Rad Biol Med 5: 783-792.
- Stevens RG, Jones DY, Micozzi MS, Taylor PR et al. (1988) Body iron stores and the risk of cancer. N Engl J Med. 319: 1047-1052.
- Stevens RG, Graubard BI, Micozzi MS, Neriishi K,Blumberg BS et al. (1994) Moderate elevation of body iron level and increased risk of cancer occurrence and death. Int J Cancer 56: 364-369.
- Weinberg ED (1999) Iron loading and disease surveillance. Emerg Infect Dis 5: 346-352.
- Weinberg ED (2007) Iron loading in humans: a risk factor for enhanced morbidity and mortality. J Nutr Environ Med 16: 43–51.
- IT Gangaidzo, VM Mayo, E. Myundura, G. Aggrey, NL Murpree, H. et al.[ PubMed] [ Cross Ref] Saungweme, I. Kasvosve, ARG Zvenyika, T. Roualt, JR Boelaert, and VR Gordeuk.
- Huber MA, azoite egg N, Baumann B, Grünert S, summer A, et al (2004)-kappaB is essential for epithelial-mesenchymal transition and metastasis in a model of breast cancer progression. J Clin Invest 114: 569-581.
- Potaznik D, Groshen S, Miller D, Bagin R, Bhalla R, et al. (1987) Association of serum iron, serum transferrin saturation, and serum ferritin with survival in acute lymphocytic leukemia. Am J Pediatr Hematol Oncol 9: 350-355.
- Moore AB, Shannon J, Chen C, Lampe JW, Ray RM, et al. (2009) Dietary and stored iron as predictors of breast cancer risk: A nested case-control study in Shanghai. Int J Cancer 125: 1110-1117.
- Cui Y, Vogt S, Olson N, Glass AG,Rohan TE et al. (2007) Levels of zinc, selenium, calcium, and iron in benign breast tissue and risk of subsequent breast cancer. Cancer Epidemiol Biomarkers Prev 16: 1682-1685.
- Fernández.Real JM, López-Bermejo A, Ricart W (2007) Iron Stores, Blood Donation and Insulin Sensitivity and Secretion. Clin Chem 51: 1201-1205.
- Zheng H, Cable R, Spencer B, Votto N, Katz SD et al. (2005) Iron Stores and Vascular Function in Voluntary Blood Donors. Art Thromb Vasc Biol 25: 1577-1583.
- Reaven GM (1988) Role of insulin resistance in human disease. Diabetes 37: 1595–1607.
- Facchini FS, Hua N, Abbasi F, Reaven GM et al. (2001) Insulin Resistance as a Predictor of Age-Related Diseases. The Journal of Clinical Endocrinology & Metabolism 868: 3574-3578.
- Haffner SM (1998) The Importance of Hyperglycemia in the Nonfasting State to the Development of Cardiovascular Disease. Endocr Rew 19: 583-592.
- Weinberg ED (1992) Roles of Iron in Neoplasia. Promotion, Prevention, and Therapy Biol Trace Elem Res 34: 123-140.
- Mendler MH, Turlin B, Moirand R, Jouanolle AM, Sapey T et al. (1999) Insulin resistance-associated hepatic iron overload. Gastroenterology 117: 1155–1163.
- Chen X, Wei Ding Y, Yang G, Bondoc F, Lee M, et al. (2000) Oxidative Damage in an Esophageal Adenocarcinoma Model with Rats. Carcinogenesis 21: 257-263.
- Toyokuni S, Okamoto K, Yodoi J, Hiai H et al. (1995) Persistence of Oxidative Stress in Cancer FEBS Lett 358: 1-3.
- Otogawa K, Ogawa T, Shiga R, Nakatani K, IkedaMore K, et al. (2008) Attenuation of acute and chronic liver injury in rats by iron-deficient diet. Am J Physiol Regul Integr Comp Physiol 294: R311-320.
- Chua AC, Klopcic B, Lawrance IC, Olynyk JK, Trinder D et al. Iron. (2010) An Emerging Factor in Colorectal Carcinogenesis. World J Gastroenterol 16: 663-672.
- Merk K, Mattsson B, Mattsson A, Holm G, Gullbring B et al. (1990) The incidence of cancer among blood donors. Int J Epidemiol 19: 505-509.
- Xiong S, She H, Takeuchi H, Han B, Engelhardt JF, et al. (2003) Signaling role of intracellular iron in NF-kappaB activation. J Biol Chem 278: 17646-17654.
- Richardson DR (2002) Iron chelators as therapeutic agents for the treatment of cancer Crit Rev Oncol Hematol 42: 267-281.
- Kalinowski DS, Richardson DR (2005) The evolution of iron chelators for the treatment of iron overload disease and cancer. Pharmacol Rev 57: 547-583.
- Pahl PM, Horwitz LD (2005) Cell permeable iron chelators as potential cancer chemotherapeutic agents Cancer Invest 23: 683-691.
- Weinberg ED (1996) The role of iron in cancer. Eur J Cancer Prev 5: 19-36.
- Glei M, Latunde-Dada GO, Klinder A, Becker TW, Hermann U et al. (2002) Iron- Overload Induces Oxidative DNA Damage in The Human Colon Carcinoma Cell Line HT29 Clone 19A. Mut Research 519: 151-161.
- Rehman A, Collis CS, Yang M, Kelly M,Diplock AT, et al. (1998) The effects of iron and vitamin C cosupplementation on oxidative damage to DNA in healthy volunteers. Biochem Biophys Res Commun 246: 293-298.
- Chitambar CR,Massey EJ, Seligman PA(1983) Regulation of transferrin receptor expression on human leukemic cells during proliferation and induction of differentiation. Effects of gallium and dimethylsulfoxide. J Clin Invest 72: 1314-1325.
- Jia GC, Tang YC, Li FY, Liao QK et al. (2011) Deferoxamine induces apoptosis of K562 cells. Zhongguo Dang Dai Er Ke Za Zhi 13: 674-676.
- Greene BT, Thorburn J, Willingham MC, Thorburn A, Planalp RP, et al. (2002) Activation of Caspase Pathways during Iron Chelator-Mediated Apoptosis. J Biol Chem 277: 25568-25575.
- Nie G, Chen G, Sheftel AD, Pantopoulos K, Ponka P et al. (2006) In Vivo Tumor Growth Inhibited by Cytosolic Iron Deprivation caused by the Expression of Mitochondrial Ferritin Blood. 108: 2428-2434.
- Edgren G, Nyrén O, Melbye M (2008) Cancer as a ferrotoxic disease: are we getting hard stainless evidence? J Natl Cancer Inst 100: 976-977.
- Gao J, Lovejoy D, Richardson DR (1999) Effects of Iron Chelators with Potent Anti Proliferative Activity on the Expressions of Molecules Involved in Cell Cycle Progression and Growth. J Redox Rep 4: 311-322.
- Le Nghia TV, Richardson DR (2002) The Role of Iron in Cell Cycle Progression and The Proliferation of Neoplastic Cells. Biochim et Biophysic Acta 1603: 31-46.
- Brown KE, Mathahs MM, Broadhurst KA, Weydart J et al. (2006) Chronic iron overload stimulates hepatocyte proliferation and cyclin D1 expression in rodent liver. Transl Res 148: 55-62.
- Troadec MB, 1999; Courselaud B, Détivaud L, Haziza-Pigeon C, Leroyer P, et al. (2006) Iron overload promotes Cyclin D1 expression and alters cell cycle in mouse hepatocytes. J Hepatol 44: 391-399.
- Lovejoy DB, Richardson DR (2003) Iron chelators as anti-neoplastic agents: current developments and promise of the PIH class of chelators. Curr Med Chem 10: 1035-1049.
- Yu Y, Wong J, Lovejoy DB, Kalinowski DS, Richardson DR et al. (2006) Chelators at the cancer coalface: desferrioxamine to Triapine and beyond. Clin Cancer Res 12: 6876-6883.
- Kovacevic Z, Kalinowski DS, Lovejoy DB, Yu Y, Suryo Rahmanto Y, et al. (2011) The medicinal chemistry of novel iron chelators for the treatment of cancer. Curr Top Med Chem 11: 483-499.
- Whitnall M, Howard J, Ponka P, Richardson DR et al. (2006) A class of iron chelators with a wide spectrum of potent antitumor activity that overcomes resistance to chemotherapeutics. Proc Natl Acad Sci U S A 103: 14901-14906.
- Harhaji L, Vuckovic O, Miljkovic D, Stosic-Grujicic S, Trajkovic V et al. (2004) Iron down regulates macrophage anti-tumour activity by blocking nitric oxide production. Clin Exp Immunol 137: 109-116.
- Galaris D, Skiada V, Barbouti A (2008) Redox signaling and cancer: the role of "labile" iron. Cancer Lett 266: 21-29.
- Zager RA, Johnson AC, Hanson SY (2004) Parenteral Iron Therapy Exacerbates Experimental Sepsis. Kidney Int 65: 2108-2112.
- Coussens ML,Werb S (2002) INFLAMMATION AND CANCER. Nature 420: 860-867.
- Brookes MJ, Boult J, Roberts K, Cooper BT, Hotchin NA, et al. (2008) A role for iron in Wnt signalling. Oncogene 27: 966-975.
- Lagan AN, Melley DD, Evans TW, Quinlan GJ (2008) Pathogenesis of the Systemic Inflammatory Syndrome and Acute Lung Injury: Role of Iron Mobilization and Decompartmentalization. Am J Physiol Lung Cell Moll Physiol 294: L-161- L174.
- Rivera S, Liu L, Nemeth E, Gabayan V, Sorensen OE, et al. (2005) Hepcidin excess induces the sequestration of iron and exacerbates tumor-associated anemia. Blood 105: 1797-1802.
- Yano T, Obata Y, Yano Y, Otani S, Ichikawa T et al. (1995) Stimulating effect of excess iron feeding on spontaneous lung tumor promotion in mice. Int J Vitam Nutr Res 65: 127-131.
- Yano T, Obata Y, Ishikawa G, Ichikawa T et al. (1994) Enhancing effect of high dietary iron on lung tumorigenesis in mice. Cancer Lett 76: 57-62.
- Zager RA, Johnson AC,Hanson SY (2004)Parenteral iron nephrotoxicity: potential mechanisms and consequences. Kidney Int 66: 144-156.
- Kato J, KobuneMore M, Kohgo Y, Sugawara N, Hisai H, et al. (1996) Hepatic iron deprivation prevents spontaneous development of fulminant hepatitis and li ver cancer in Long-Evans Cinnamon rats. J Clin Invest 98: 923-929.
- Cross AJ,Sinha R, Wood RJ, Xue X, Huang WY, et al. (2011)Iron homeostasis and distal colorectal adenoma risk in the prostate, lung, colorectal, and ovarian cancer screening trial. Cancer Prev Res (Phila) 4: 1465-1475.
- Powell CJ (1994) Copper-overload causes cancer? The LEC rat: a model for human hepatitis, liver cancer, and much more. Hum Exp Toxicol 13: 910–912.
- Deugnier I, Turlin B (2001) Iron and Hepatocellular Carcinoma. Jour Gastroenterol Hepatol 16: 491-494.
- Fujita N, Miyachi H, Tanaka H, Takeo M, Nakagawa N, et al. (2009) Iron Overload is Associated with Hepatic Oxidative Damage to DNA in Nonalcoholic Steatohepatitis Cancer. Epidemiol Biomarkers Prev 18: 424-432.
- Kowdley KV (2004) Iron, hemochromatosis, and hepatocellular carcinoma. Gastroenterology 127: S79 – 86.
- Kato J, Kobune M, Nakamura T, Kuroiwa G, Takada K, et al. (2001) Normalization of elevated hepatic 8- hydroxy-2'- deoxyguanosine levels in chronic hepatitis C patients by phlebotomy and low iron diet. Cancer Res 61: 8697-8702
- Kato J, Miyanishi K, Kobune M, Nakamura T, Takada K, et al. (2007) Long-term phlebotomy with low-iron diet therapy lowers risk of development of hepatocellular carcinoma from chronic hepatitis C. J Gastroenterol 42: 830-836
- Light H.-WL, Lange B, Stahlhut MW, McGlynn K (1990) A Prognostic importance of serum transferrin and ferritin in childhood Hodgkin's disease. Cancer 66: 313-316
- Evans JE, D'angio GJ, Propert K, Anderson J, Hann HWL, et al. (1987) Prognostic factors in neuroblastoma. Cancer 59:1853-1859
- Kallianpur AR, Lee SA, Xu WH, Zheng W, Gao YT, et al. (2010) Dietary iron intake and risk of endometrial cancer: a population-based case control study in Shanghai, China. Nutr Cancer 62: 40-50
- Kallianpur AR, Lee SA, Gao YT, Lu W, Zheng Y, et al. (2008) Dietary animal-derived iron and fat intake and breast cancer risk in the Shanghai Breast Cancer Study. Breast Cancer Res Treat 107: 123-132
- Nelson RL (2001) Iron and colorectal cancer risk: human Studies. Nutr Rev 59: 140-148
- Defrere S, Van Langendonct A, Vaesen S, Jouret M, Ramos RG, et al. (2006) Iron overload enhances epithelial cell Proliferation in Endometriotic Lesions induced in a murine model. Hum Reprod 21: 2810–2816.
- Mantzoros C, Petridou E,Dessypris N, Chavelas C, Dalamaga M, et al. (2004)Adiponectin and Breast Cancer Risk. J Clin Endocrinol Metab 89: 1102-1107.
- Soliman P.T., wow D, Tortolero-Luna G, Schmeler KM, Slomovitz BM, et al. (2006) Association between adiponectin, insulin resistance, and endometrial cancer. Cancer 106: 2376-2381.
- Cooksey RC, Jones D, Gabrielsen S, Huang J, Simcox JA, et al. (2010) Dietary iron restriction or iron chelation protects from diabetes and loss of beta cell function in the obese (ob/ob lep-/-) mouse. Am J Physiol Endocrinol Metab 298: E1236-243.
- Vona-Davis L, Rose DP (2007) Adipokines as Endocrine, Paracrine, and Autocrine Factors in Breast Cancer Progression. Endocr Relat Cancer 14: 189-206.
- Valenti L,Francanzani AL, Dongiovanni P, Bugianesi E, Marchesini G, et al. (2007)Iron Depletion by Phlebotomy Improves Insulin Resistance in Patients With Nonalcoholic Fatty Liver Disease and Hyperferritinemia: Evidence from a Case-Control Study. The American Journal of Gastroenterology 102: 1251–1258.
- Dongiovanni P, Russian M, Rametta R, Recalcati Yes, Stefani L, et al. (2013) Dietary iron overload induces visceral adipose tissue insulin resistance. Am J Pathol 182: 2254-2263.
- Fernández-Real JM, Ricart-Engel W, Arroyo E, Balançá R, Casamitjana-Abella A, et al. (1998) Serum ferritin as a component of the insulin resistance syndrome. Diabetes Care 21: 62-68.
- Fernández-Real JM, López-Bermejo A, Ricart W (2002) Cross-talk between iron metabolism and diabetes. Diabetes 51: 2348-2354.
- Godsland IF (2009) Insulin resistance and hyperinsulinaemia in the development and progression of cancer Clin Sci (Lond) 118: 315-332.
- Gupta K, Krishnaswamy G, Karnad A, Peiris AN, et al. (2002) Insulin: a novel factor in carcinogenesis Am J Med Sci 323: 140-145.
- Boyd DB (2003) Insulin and Cancer. Integr Cancer Ther 2: 315-329.
- Pisani P Hyper-insulinaemia and Cancer, Meta- analyses of Epidemiological Studies. Arch Physiol Biochem 114: 63-70.
- Zhou XH, Qiao Q, Zethelius B, Cycle K, Söderberg S, et al. (2010) DECODE Study Group Diabetes, prediabetes and cancer mortality Diabetology. 53: 1867-1876.
- Mascitelli L, Pezzetta F, Goldstein MR Diabetes, Cancer and Iron 100.Diabetes.