Mechanisms of respiration in aquatic Arthropods
Hassan Vatandoost1*,2
1Professor, Department of Medical Entomology & Vector Control, School of Public Health, Tehran University of Medical Sciences, P.O. Box:6446-14155, Tehran, Iran.
2Department of chemical pollutants and Pesticides, Institute for Environmental Research, Tehran University of Medical Sciences, Tehran, Iran.
*Corresponding Author:Hassan Vatandoost, Professor, Department of Medical Entomology & Vector Control, School of Public Health, Tehran University of Medical Sciences, P.O. Box:6446-14155, Tehran, Iran, Tel: +98 21 42933225; Fax: +98 21 42933225; E-mail: hvatandoost1@yahoo.com
Citation: Hassan Vatandoost (2023)Mechanisms of respiration in aquatic Arthropods. SciEnvironm 6: 179.
Received: April 18, 2023; Accepted: May 10, 2023; Published: May 15, 2023.
There are no oxygen-carrying compounds in the blood stream of arthropods, therefore they transfer the oxygen in specific conditions. The waterproof cuticle is an important factor. For some arthropods the environment with humidity is appropriate for their life. The cuticle mechanism of arthropods is water permeability. The required oxygen is diffusing from the cuticle However, most insects have a special respiratory system comprising a system of internal tubes, called trachea, which branch and rebranch. Very fine branches, tracheoles, penetrate individual cells. Spiral stiffening is the function of trachea. The act as vacuum cleaner hose. The trachea prevent collapse. The opening spiracles allow the air to perpetrate into the body. In their thorax, normally two pairs of spiracles are present. Their abdomen also has 8 pairs. All the spiracles are located laterally in the body. Oxygen transfer from spiracles to tracheae and from tracheole to cells (Fig.1). Rate of gas diffusion is determined by differences in concentration at each end of the system. Molecular weight of the gas. So, most of the path for oxygen to diffuse from the spiracle to the cell is through the air-filled tracheal tubes.Insects need to avoid water loss through their spiracles and to prevent contamination by dust. Spiracles are therefore usually equipped with opening and closing devices and filtering lobes or hairs in an atrium before the beginning of the trachae. Internal gills could be observed in some arthropods. They are part of the tracheal system. Mechanical ventilation can be observed. In some species of Hymenoptera, there are specific mechanism for breathing. They pump the air by extending the abdomens. Many aquatic insects must return periodically to the surface of water to breathe. Mosquito larvae have tubes connecting to the air at the water surface. There are hydrofuge hairs (water resistant) associated with the spiracles, to break the surface tension of the water and to keep water out when the insect is underwater. Stone fly, dragonfly and mayfly nymphs have tracheal gills whereby oxygen diffuses from the water to fine, surface trachae. Small internal parasites may diffuse oxygen across their cuticle. Others may have spiracles at the end of siphons which open into the trachae of their host or penetrate the skin of their host into the air. The cabbage looper was parasitized by Tachinid flies. The dark spots are holes for the maggot's breathing tubes (Fig.2).
Figure 1: Spiracle in insects.
Figure 2: Maggot breathing tubes in the body of host.

Mayfly larvae are very interesting for a study of the insect respiratory system. Mayfly larvae are fully aquatic. Trachea are connected to the gill-like form. In this species they have along the body as the long strands on the right. They branch into finer tubes so all the parts of the body can be reached (Fig.3).
Figure 3: Respiratory system in mayfly nymph.

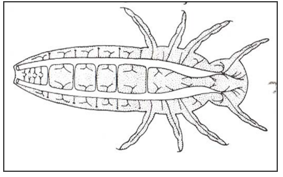
Different types of respiratory system: Open polypneustic tracheal system (Fig. 4). polypneustic tracheal system with air-sacs for ventilation (Fig.5). Oligopneustic tracheal system, terminal spiracles alone functional (Fig.6).
Figure 4: Open polypneustic tracheal system.
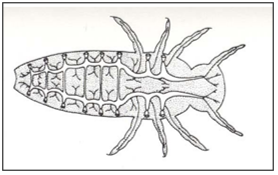
Figure 5: Polypneustic tracheal system with air-sacs for ventilation.
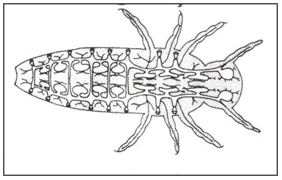
Figure 6: Oligopneustic tracheal system, terminal spiracles alone functional.
In atmospheric breather (mosquito larvae and pupae), at the end of spiracles there are hydrofuge, water repellent, cuticle or hair. Lobes seals the spiracle opening. Maintenance of air bubble that covers the opening (Fig.7). In plant breather (spiracles modified to pierce submerged portions of aquatic plants and the parenchyma air stores (Fig.8).
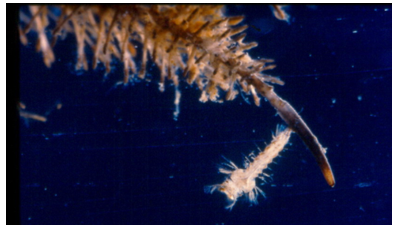
Figure 7: Rat-tailed maggot siphon in Syrphidae family.
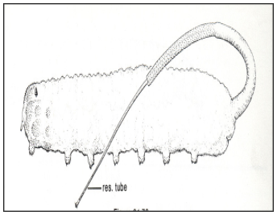
Figure 8: Larvae of Mansonia mosquito (plant breather).
In closed spiracle there are lack direct communication with the outside and rely on gaseous diffusion through the cuticle for respiratory exchange (apneustic) (Fig.9). Closed tracheal system with abdominal tracheal gills (Fig.10). Closed tracheal system with rectal tracheal gills (internal gills) Dragonfly (Fig.11).
Figure 9: Closed tracheal system, cutaneous respiration only.
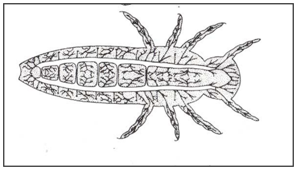
Figure 10: Closed tracheal system with abdominal tracheal gills.
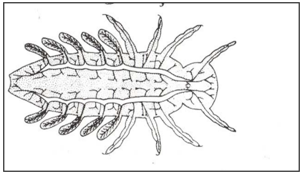
Figure 11: Closed tracheal system with rectal tracheal gills (internal gills) Dragonfly.
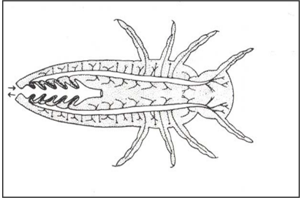
Head gills and Cervical gills could be observed in Plecoptera family (Fig.12). Lateral gill is shown in Fig.13.
Figure 12: Head gills, Cervical gills. Plecoptera.
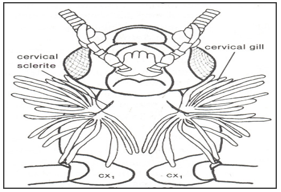
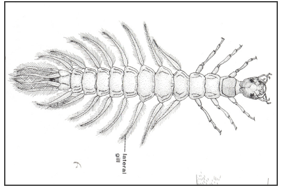
Figure 13: Lateral gills.
Ventral air bubble, which also serves as a temporary air store (physical gill, compressible gill), backswimmer, Neoplea. Water beetles take a bubble of air with them underwater. Their spiracles open into the bubble. As the oxygen in the bubble is used up, the oxygen pressure in the bubble drops to that of the surrounding water so that more oxygen diffuses into the bubble from the water. In this way the bubble remains usable for a long time. (Fig.14).
Figure 14: Temporary air store (physical gill, compressible gill).
Figure 15: Hydrofuge cuticular.
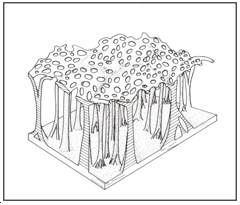
Hydrofuge cuticular network found in the spiracular gill of the tipulid Dicranomyia sp (Fig. 15).
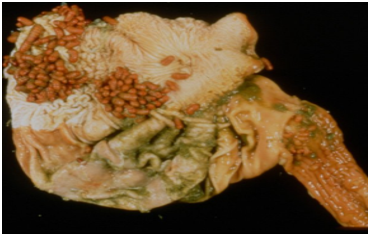
Respiratory pigment. Homoglobin in bloodworm absorb oxygen. It is found in Chironomidae, Notonectidae and Gastrophilidae families (Fig.16,17,18). Bloodworms, (midge larvae) live in the mud of stagnant pools where there is very little oxygen. These insect larvae are one of the few which have haemoglobin in their blood and this haemoglobin is more efficient than humans.
Figure 16: Chironomidae larva with haemoglobin in the blood.
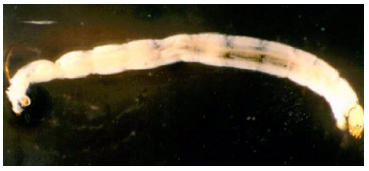
Figure 17: Notonectidae family, Backs swimmer.

Figure 18: Gastrophilidae Larvae (in stomach of horse).
References
- Bradley TJ (2000) The discontinuous gas exchange cycle in insects may serve to reduce oxygen supply to the tissues. Am Zool 40: 952.
- Buck J (1958) Cyclic CO2 release in insects. IV. A theory of mechanism. Biol. Bull 114: 118-140.
- Chown SL, Gibbs AG, Hetz SK, Klok CJ, Lighton JRB et al. (2006) Discontinuous gas exchange in insects: a clarification of hypotheses and approaches. Physiol Biochem Zool 79: 333-343.
- Chown SL,Holter P (2000)Discontinuous gas exchange cycles in Aphodius fossor (Scarabaeidae): a test of hypotheses concerning origins and mechanisms. J Exp Biol 203: 397-403.
- Gibbs AG,Johnson RA (2004) The role of the discontinuous gas exchange in insects: the chthonic hypothesis does not hold water. J Exp Biol 207: 3477-3482.
- Harrison JF, Hadley NF, Quinlan MC (1995) Acid-base status and spiracular control during discontinuous ventilation in grasshoppers. J Exp Biol 198: 1755-1763.
- Hetz SK, Bradley TJ (2005) Insects breathe discontinuously to avoid oxygen toxicity. Nature 433: 516-519.
- Hetz SL, Wasserthal LT, Heermann S, Kaden H, Oelssner W et al. (1994) Direct oxygen measurements in the tracheal system of lepidopterous pupae using miniaturized amperometric sensors. Bioelectrochem Bioenerg 33: 165-170.
- Levy RJ, Schneiderman HA (1966) Discontinuous respiration in insects. II. the direct measurement and significance of changes in tracheal gas composition during the respiratory cycle of silkworm pupae. J Insect Physiol 12: 83-104.
- Lighton, JRB, Berrigan D (1995) Questioning paradigms: caste-specific ventilation in harvester ants, Messor pergandei and M. julianus (Hymenopetera: Formicidae). J Exp Biol 198: 521-530.
- Lighton JRB (1996) Discontinuous gas exchange in insects. Annu Rev Entomol 41: 309-324.
- Lighton JRB (1998) Notes from the underground: towards the ultimate hypotheses of cyclic, discontinuous gas–exchange in tracheate arthropods. Am Zool 38: 483-491.
- Nation JL Insect Physiology and Biochemistry. (2002) CRC, Boca Raton, FL.
- Slama K A (1988) New look at insect respiration. Biol Bull 175: 289-300.
- Williams AE, Rose MR, Bradley TJ (1997) CO2 release patterns in Drosophila melanogaster: the effect of selection for desiccation resistance. J Exp Biol 200: 615- 624.