The Genetic Base for Drug Addiction
Srwsh N Qadr1*
1 Kscien Organization for Scientific Research, Sulaimani, Kurdistan, Iraq.
*Corresponding Author:Srwsh N. Qadr, Kscien Organization for Scientific Research, Sulaimani, Kurdistan, Iraq
Citation: Srwsh N Qadr (2023) The Genetic Base for Drug Addiction. Addict drug sensitiz 4: 121.
Received: April 11, 2023; Accepted: April 26, 2023; Published: April 29, 2023.
Copyright: © 2023 Srwsh N. Qadr, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Abstract
Objective: There are many evidences about relationship between eating behavior and drug addiction. A number of susceptibility loci that point to shared higher order genetic pathways underling addiction were found in genetic studies. This study assumed that a genome-wide association study (GWAS) of food addiction would produce important enrichment in genes and pathways related to addiction.
Methods: This study done among 314 women of European ancestry, by using a GWAS of food addiction, which is determined by the modified Yale Food Addiction Scale (mYFAS). Results for enrichment of single nucleotide polymorphisms (SNPs) (n 5 44), genes (n 5 238), and pathways (n 5 11) involved in drug addiction were tested.
Results: Two loci met GW-significance (P < 2.5 10 -8) with no obvious roles in eating behavior, they are mapping to 17q21.31 and 11q13.4. GW results were significantly enriched for gene members of the MAPK signaling pathway (P = 0.02). After adjustment for multiple testing, candidate SNP or gene for drug addiction was not linked with food addiction.
Conclusions: limited support was delivered for shared genetic underpinnings of drug addiction and food addiction, although the GWAS of mYFAS, need further investigation and follow up.
Keywords: Gene; Food; Addiction.
Introduction
In behavioral and neurobiological investigation evidences of association between addiction and feeding behavior is accumulated [1]. Viewing obesity as a neurobehavioral disorder as a result of interaction between a vulnerable brain and environment is indicative of models of drug addiction [2]. Evidences supporting a behavioral component of obesity was yielded on genome-wide association studies (GWAS) of adiposity and follow up studies of confirmed loci [3-5]. Early candidate and more recent GWAS of addictive behaviors have given rise to a set of susceptible loci indicating common genetic pathways of higher order underlying addictive behavior [6-8], Consequently, the opportunity to clarify whether specific genetic influences on drug dependency generalize addictive eating behaviors has been provided.
The Yale Food Addiction Scale (YFAS) is a psychometric tool for assessing food addiction in individuals based on the Diagnostic and Statistical Manual for Mental Disorders (DSM)-IV codes for substance dependence criteria [9]. Previously, the YFAS was associated with episodes of binge-eating, emotional eating, hedonic eating, impulsivity, and craving for food and snack as well as neural response patterns involved in other addictive disorders [10]. Recently, food addiction was assessed in women participating in the Nurses ' Health Study (NHS) and Nurses ' Health Study II (NHS2) using a modified version of the YFAS with similar psychometric properties compared to the original YFAS [11]. Our goal was to conduct the first comprehensive genetic analysis of food addiction in a population among the subset of these women who have GW scan data. If food addiction shares a molecular pathophysiology with classical addictions as captured by mYFAS, we hypothesized that a GWAS of food addiction would yield substantial enrichment in addiction-related genes and pathways.
Methods
Study Populations: The NHS was founded in 1976 with 121,700 registered female nurses aged 30 - 55 and residing in 11 U.S. states [12]. The NHS2 cohort was set up in 1989 with 116,609 female nurses aged 25-44 and residing in 14 U.S. states [13]. Women in both cohorts received mailed questionnaires on medical history and lifestyle characteristics every 2 years [12,13]. Brigham and Women's Hospital's Institutional review board approved the study protocols.
Measures: Food addiction assessment and covariates. The YFAS consists of 25 questionnaire items used to evaluate diagnostic criteria for food addiction and provides both a number of symptoms of food addiction and a diagnosis of food addiction as scoring options [9,14]. We have recently created and validated a modified YFAS (mYFAS) for use in large epidemiological cohorts by adjusting the original YFAS to a core of nine questionnaire items with one question from each of the seven diagnostic criteria plus two individual items assessing the existence of clinically significant impairment and distress (Supporting Information, Table S1). The frequency threshold from the original YFAS was used for each of the diagnostic criteria and all questions were summed up for a total score of 0 to 7 (mYscore). MYFAS's reliability, convergence, discriminatory and incremental validity were reported in detail elsewhere [11]. Briefly, in a study sample whose data were previously reported in the YFAS validation, mYscore's internal consistency was sufficient and identical to that of the full YFAS version (Kuder-Richardson α = 0.75). Also similar was the convergent and discriminating validity of mYscore and the original YFAS. Compared to the original YFAS diagnostic version and YFAS symptom count, the mYFAS diagnostic version and mYscore were also significantly associated with binge-eating scores above and beyond other eating pathology measures [11].
In 2008, the NHS administered the mYFAS questionnaire, at which time the participants were 62-87 years of age. In 2009, when participants were aged 45-64 years, mYFAS questionnaires were administered in the NHS2. The response rates for both cohorts exceeded 80% and those who responded to food addiction issues were not substantially different from those who did not have BMI or smoking status [11]. In 2012, a subset of NHS2 respondents were invited to complete the full YFAS and a strong correlation was observed in scores (Pearson's r = 0.98) and internal consistency (Cronbach's coefficient α = 0.84) [11].
We examined two features of food addiction for the current GW analysis: i) food addiction symptoms (mYscore) modeled as a continuous or dichotomous (mYscore 3) feature and ii) clinically significant impairment and distress (Yclinical) presence / absence. MYscore and Yclinical derive from a number of different questions as detailed in Supporting Information, Table S1. The combined presence of more than 3 symptoms of food addiction (mYscore) and significant impairment or distress (Yclinical) are proposed food addiction diagnostic criteria (mYdiag) [9,11].
All covariates were collected through a self-administered questionnaire and at the same time as measures for food addiction. BMI (kg / m2) was derived from the self-reported weight and height reported in our cohorts with high precision [15].
 Genotyping, control of quality, and imputation. Between 1989 and 1990, Blood was collected from 32,826 NHS members and from 29,611 NHS2 members from 1996 to 1999. White blood cells extracted DNA. Women contributing to the recent genetic testing were those previously selected in nested case-control studies for a variety of chronic diseases for independent GWAS. (Supporting Information, Table 1). We pooled genotyped samples on the same platforms to maximize efficiency and power, resulting in three data sets called Affy (NHS), Illumina (NHS, NHS2), and Omni (NHS). Thorough mechanisms and quality assurance of these genetic data sets (S Lindstrom, S Loomis, Chen, unpublished data) have been reported and relevant descriptive and quality control (QC) data are provided in the Supporting Information. Table S1. Any samples with significant genetic resemblance to non-European samples of reference were excluded. MACH (v.1.0.18.c) and Minimac (v.2012-08-15) were used for each of the three data sets to impute about 31 million single-nucleotide polymorphisms (SNPs) based on the reference panel of 1000 G v3 ALL.
Genotyping, control of quality, and imputation. Between 1989 and 1990, Blood was collected from 32,826 NHS members and from 29,611 NHS2 members from 1996 to 1999. White blood cells extracted DNA. Women contributing to the recent genetic testing were those previously selected in nested case-control studies for a variety of chronic diseases for independent GWAS. (Supporting Information, Table 1). We pooled genotyped samples on the same platforms to maximize efficiency and power, resulting in three data sets called Affy (NHS), Illumina (NHS, NHS2), and Omni (NHS). Thorough mechanisms and quality assurance of these genetic data sets (S Lindstrom, S Loomis, Chen, unpublished data) have been reported and relevant descriptive and quality control (QC) data are provided in the Supporting Information. Table S1. Any samples with significant genetic resemblance to non-European samples of reference were excluded. MACH (v.1.0.18.c) and Minimac (v.2012-08-15) were used for each of the three data sets to impute about 31 million single-nucleotide polymorphisms (SNPs) based on the reference panel of 1000 G v3 ALL.
Statistical analysis: GW analysis of mYFAS. For each cohort of women, each genetic data set was examined separately and meta-analysis combined the results. There are NHS and NHS2 data within the Illumina data set. Four data sets were therefore investigated: Affy-NHS (N 5 3298), Illumina-NHS (N 5 2690), Omni-NHS (N 5 2520), and Illumina-NHS2 (N 5 806). Based on linear (mYscore) or logistic (mYscore 3, Yclinical) regression under an additive genetic model and age adjustment, BMI, initial case-control data set, and four main components of population substructure, we performed GWA testing for each trait over 31 million SNPs (expressed as allele dosage). Before meta-analysis (Supporting Information, Table S1), SNPs with minor allele frequency (MAF) < 0.3 or with low imputation quality scores (MACH’s Rsq<0.3) were removed. We removed BMI from the model in secondary analyzes or adjusted further for smoking status (current, past, never).
For both features of food addiction (mYscore, mYclinical), GW meta-analysis was performed using a model of fixed effects and inverse weighting with a single correction of genomic control (GC) as per METAL [16]. The heterogeneity of the set of inter data was investigated using the I2 statistics [17]. Top loci associated with each trait were retained and formally presented if i) SNPs passed QC filters across all four data sets and ii) effect direction across all data sets was consistent.
GW-significance was defined as P < 2.5 × 10-8, the traditional threshold (P < 5 × 10-8) [18] for the number of independent characteristics corrected. Nominally significant loci were tabulated if all data sets and features of food addiction were consistent in the direction of effects. Top loci in the NHS (the larger of the two contributing cohorts) were examined for associations with mYdiag, BMI, and smoking. Our full results for BMI associations were also investigated on the basis of a published large-scale GWAS [3,19].
Candidate SNP, gene-set, and pathway analysis. For evidence of overlap with SNPs, genes and pathways involved in drug addiction, we interviewed summary-level results from our mYscore and Yclinical GWAS. "Addiction SNPs" included 44 common (r2 < 0.8) independent (MAF > 0.01) SNPs with at least nominal significance (300 kb) of each SNP addiction. A total of 238 genes were regarded as genes of addiction (Supporting Information). Gene-based analyzes of candidates were carried out using VErsatile Gene-based Association Study (VEGAS) [20]. We applied P < 0.001 [0.05/44 (number of SNPs tested) ] and P < 2.1 × 10-4 [0.05/238 (number of genes tested) ] Bonferroni-corrected thresholds for SNP-level and gene-level meaning respectively. Additionally, multi-SNP linear kernel tests were used to evaluate the relationship between the 44 addiction SNPs and food addiction [21]. These linear models allow multiple SNP associations to be tested concurrently with one test and do not necessitate risk allele direction pre-specification.
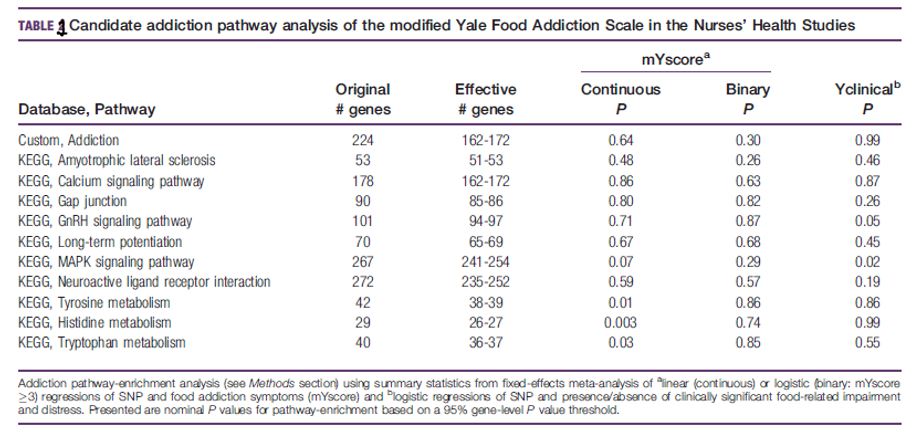
Meta-Analysis Gene-set VariaNT Associations (MAGENTA) enrichment [22] has been used to test our GW mYFAS results for enrichment of the addiction pathway. We observed consistent enrichment of genes related to the Kegg pathways in preliminary pathway analyzes of our addiction genes (defined above): (1) "tyrosine metabolism," (2) "amyotrophic lateral sclerosis," (3) "calcium signaling pathway," (4) "neuroactive ligand receptor interaction," (5) "tryptophan metabolism," (6) "long-term potentialation" and (7) "histidine metabolism," (Supporting Information,).
Two of all these pathways intersected with those reported by Li et al. (8), who also reported over-enrichment of genes related to "GnRH signaling pathway," (8) "MAPK signaling pathway," (9) and "Gap junctions" (10). We also created an (11) addiction gene set which included the above-defined addiction genes (excluding 14 genes: hypothetical or pseudogenic or non-MAGENTA-annotated genes).
Taken together, specifically 11 gene sets or pathways were tested and the nominal meaning threshold of 0.05 was applied to MAGENTA. Exploration was supplemented with hypothesis testing: results from seven databases were tested against 3,218 pathways. For each pathway, the enrichment of highly ranked gene scores in meta-analysis above the 95th percentile of all gene scores was evaluated compared to 10,000 randomly sampled gene sets [22].
Results
GWAS of mYFAS. Two loci met GW-significance: General features of the 9,314 women included in the present analysis are shown in Supporting Information Table S1. The mean mYscore for NHS and NHS2 (standard deviation, SD) was 0.60 (1.05) and 1.04 (1.6), respectively. Yclinical and mYdiag prevalence in the NHS was 4.9 percent and 2.6 percent. The NHS2's corresponding prevalence was 11.2% and 8.7%. In both cohorts, MYscore was correlated with BMI (NHS Pearson’s r = 0.27, NHS2 Pearson’s r = 0.42; P < 0.001). These results are similar to those reported in the full NHS and NHS2 cohorts, indicating an increased presence of food addiction in the younger cohort [11].
Characteristics of NHS and NHS2: criteria for food addiction characteristics in GWAS (Table 1, Supporting Information, Figures S1-S3). MYscore (P = 2.0× 10-8 ) and Yclinical (P = 6.4 × 10-4) were associated with SNPs at 17q21.31 mapping to the intronic region of PRKCA. In the NHS, a positive mYdiag (P = 3.4 × 10-5) was also associated with the variant of the 17q21.31 index SNP rs74902201 with higher mYscore. MYscore was significantly associated with SNPs at 11q13.4 mapping to the NTM intronic region, but not with Yclinical.
The variant of the 11q13.4 index SNP rs75038630 associated with higher mYscore was also associated with a positive mYdiag (P = 1.7 × 10-4) but a lower BMI (P = 0.02) in the NHS. Between an intergenic 6q22.32 locus near CENPW and mYscore (P = 3.1× 10-8) a borderline GW significant association was observed. In the NHS, a positive mYdiag (P = 1.4× 10-5) was associated with the variant of the 6q22.32 index SNP rs139878170 associated with higher mYscore. Removing BMI from the regression model reinforced the association between rs139878170 and mYscore (P = 6.9× 10-9,). Loci (or proxies) reported in Table 1 were not linked to BMI by a published BMI large-scale GWAS (3). Of the 32 BMI loci identified in the latter, mYscore was associated with rs1558902 (FTO, P = 0.04), rs206936 (NUDT3, P = 0.01), and rs10150332 (NRXN3, P = 0.05).
Candidate addiction SNPs: No association of candidate SNPs and features of food addiction met our prescribed threshold of significance (P < 0.001). Only a nominally significant association between the intergenic SNP rs1868152 [ previously associated with illicit drug use [23] ] for mYscore and Yclinical (P < 0.004, Supporting Information, Table 1 was observed among the 44 SNPs previously associated with addiction traits in GWAS. Results from linear multi-SNP kernel tests across Affy-NHS, Omni-NHS, Illumina-NHS, and Illumina-NHS2 were not consistent. Tests in NHS-Affy (P = 0.03) and Yclinical in NHS2- illumina (P = 0.01) were only significant for dichotomically modeled mYscore. There were no significant tests carried out on the combined data sets (P > 0.07).
Candidate addiction pathways: Our preset significance threshold (P < 2.1 × 10-4) was not met by a candidate gene-based test for food addiction trait associations (Supporting Information, Table 1). The most statistically significant gene was LOC100130673 (P < 5.0 × 10-4 for mYscore), a chromosome 7 pseudogene selected for proximity to SNP (rs215614) smoking behavior. In this pseudogene (rs61436781, P < 9.7 × 10-6), the latter is 135 kb away from our top SNP and is not in LD (r2 < 0.2). Four addiction genes for both mYscore and Yclinical, including HOMER1, ZHX2, DRD2, and SURF6, were nominally significant (P < 0.05).
Candidate addiction genes: Yclinical results for MAPK signaling pathway genes (P = 0.02, Table 1) were significantly enriched and this same set of genes reached borderline meaning for mYscore (P = 0.07). For the continuously modeling mYscore (P < 0.03), enrichment for tyrosine, histidine, and tryptophan metabolism genes was observed. Our mYscore and Yclinical results for gene members of our custom addiction gene set have not been significantly enriched (Table 1). An exploratory GW pathway analysis of food addiction resulted in significant gene enrichment in the binding pathway [ mYscore (binary), FDR 5 0.003] for GO interleukin-1 receptor (IL1R). Also among the top pathways were larger but similar gene sets of Ingenuity (IL-10 signaling) and Biocarta (IL1R pathway).
Discussion
An ongoing debate of growing scientific interest is the concept of "food addiction" (or "eating addiction"). The good evidence supporting this condition is the overlapping neurobiological systems reportedly detected in experimental and clinical models by both abuse drugs and highly palatable foods [1]. In this research, we investigated whether food addiction genetic determinants overlap with drug addictions. To this end, we conducted the first mYFAS GWAS and recognized suggestive loci worthy of more follow-up, but offered limited support for shared genetics with drug addictions based on comprehensive SNP, gene and pathway analysis candidates.
In two populations of U.S. women of European ancestry we identified novel GW-significant loci in PRKCA and NTM. Each variant of the SNP index with a higher mYscore occurs in 6% of European populations and 0% -16% in non-European populations [24,25]. PRKCA encodes serine / threonine-protein kinase, which is calcium-activated, phospholipid- and diacylglycerol-dependent, and involves the regulation of numerous biological processes such as insulin signaling, inflammation, and protein kinase (MAPK) activity [26]. MAPK signals are highly involved in brain function as well as a drug addiction pathway for candidates [27]. PRKCA's SNP index resides in multi-tissue regulatory regions, notably brain enhancer regions [28], and changes a binding NRSF site [29], a transcriptional repressor of neuronal genes in non-neuronal tissues. Previously, PRKCA was associated with both BMI and asthma through linkage and follow-up analysis of SNP done by Murphy et al [30]. SNPs tested by Murphy et al. [30] were not correlated with mYFAS in this study (P > 0.32) or with BMI in GWAS [3]. NTM at 11q25 encodes neurotrimin, and it is extremely expressed in human brain tissue and closely related to opioid binding protein/cell adhesion molecule-like (OPCML), also on chromosome 11 [31]. This SNP region binds NR2C2, which serves as an important repressor of several nuclear receptor signaling pathways and is required for normal cerebellum development [32]. Significant for nominal associations in the 11q25 region were also reported for alcohol dependence (OPCML) [23], body fat distribution (OPCML) [33,34], and various other characteristics [35], but none of the TABLE 1 index SNPs in these reports was associated in the current study with food addiction. Also SNPs associated with food addiction traits was only one (rs4937665) of the top SNPs near NTM previously associated with IQ in GWAS [36]. Food addiction was not measured in any other large population-based study, so in an independent study we could not replicate our two novel and promising loci.
Our GW food addiction analysis provides limited support for shared food addiction and drug addiction pathways. MYFAS was not associated with literature informing SNPs and genes associated with addiction traits. Of the 11 pathways tested for addiction, only the MAPK signaling pathway met our significance threshold. In the published BMI gene-set-enrichment analysis, the same pathway was nominally significant (nominal GSEA P = 0.02) [3]. The gene members of this pathway mapped BDNK, NFKB1, and MAP2K5 to established BMI loci. Also members of this pathway are PRKCA and its neighboring calcium channel genes that may have triggered a degree of chance or, alternatively, provide support for our novel loci and a link between drug addiction and food addiction. Interestingly, interleukin-1 receptor binding, a substantial pathway in our global pathway analysis and also recently involved in addiction behavior, does not include PRKCA or NTM, suggesting in this study that additional and novel loci of sub-GW-significance will be discovered in future efforts.
Davis et al. [38] established a genetic risk score for dopamine signaling and a higher score (conferring high dopamine signaling) was reported among 121 overweight adults in those diagnosed with YFAS food addiction and a positive correlation with emotional eating, binge eating, and food cravings. Six SNPs near DRD2, SLC6A3, and COMT were included in their score. In this study, four of the six SNPs (rs1800497, rs6277, rs12364283, rs4680) were genotyped / imputed. There were no high-quality proxies available for the remaining two SNPs. Rs1800497 was associated with and in the expected direction with Yclinical (P = 0.04). Rs12364283 was associated with mYscore on a dichotomous model (P = 0.03), but the effect direction was contrary to that expected. All three genes were "candidate genes of addiction" examined in this study, but none met our criteria of significance. Study population differences could explain the discrepancies between this study and Davis et al. study [38].
GWAS of BMI identified several loci, some of which are involved in hedonic and not homeostatic obesity pathways [3-5]. BMI is not a direct measure of food addiction but is supposed to have a component of behavior that forms the basis of the hypothesis of obesity addiction. A significant positive correlation between a genetic risk score for higher BMI and smoking behaviors (smoking initiation and dose) has recently been reported by Thorgeirsson et al. [39]. Less than 10 percent of our cohort participants were current smokers, limiting our ability to perform a similar analysis. Nevertheless, in a large GWAS [3], none of our GW-significant food addiction SNPs were associated with BMI and only nominal associations were observed between 4 out of 32 validated loci of obesity and food addiction. These results are in conflict with conclusions drawn by Thorgeirsson et al, along with limited evidence of overlap with addiction pathways [39]. In the NHS, BMI was largely independent of the associations between our GW-loci and mYFAS. However, in the presence of an environment that promotes the availability of palatable foods, individuals genetically predisposed to food addiction may be more susceptible to obesity, a concept similar to that described for illicit drug addiction.
This research marks a population's first comprehensive genetic analysis of food addiction. We did not find a novel loci linked with features of food addiction that may warrant independent duplication. Furthermore, by integrating our GW food addiction results with existing genetic knowledge of drug addiction, we did not gain insight into potential genetic overlaps between food addiction and drug addiction. While results suggest that a shared determinant may be the MAPK signaling pathway, our results taken together does not suggest that the genetic underlying principles of food addiction and drug addiction are largely different.
However, given the sample size, measurement error in the assessment of food addiction, and the narrow range and low prevalence of food addiction symptomology, our study may have had modest power to detect novel loci and significant overlap with genetic variation of other addictions. In future studies, the latter could be addressed using a case-enrichment design. Furthermore, incomplete knowledge of genetic determinants of drug addiction may limit the ability to identify shared genetic determinants of food and drug addiction.
References
- Volkow ND, Wang GJ, Tomasi D, Baler RD (2013) Obesity and addiction: neurobiological overlaps. Obes Rev 14: 2-18.
- Dagher A (2009) The neurobiology of appetite: hunger as addiction. Int J Obes (Lond) 33: S30-S33.
- Speliotes EK, Willer CJ, Berndt SI, et al. (2010) Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat Genet 42: 937-948.
- Stutzmann F, Cauchi S, Durand E, et al. (2009) Common genetic variation near MC4R is associated with eating behaviour patterns in European populations. Int J Obes (Lond) 33: 373-378.
- Cornelis MC, Rimm EB, Curhan GC, et al. (2014) Obesity susceptibility loci and uncontrolled eating, emotional eating and cognitive restraint behaviors in men and women. Obesity (Silver Spring) 22: E135-E141.
- Li CY, Zhou WZ, Zhang PW, Johnson C, Wei L, et al. (2011) Meta-analysis and genome-wide interpretation of genetic susceptibility to drug addiction. BMC Genomics 12: 508.
- Uhl GR, Drgon T, Johnson C, Liu QR (2009) Addiction genetics and pleiotropic effects of common haplotypes that make polygenic contributions to vulnerability to substance dependence. J Neurogenetics 23: 272-282.
- Li CY, Mao X, Wei L (2008) Genes and (common) pathways underlying drug addiction. PLoS Comput Biol 4: e2
- Gearhardt AN, Corbin WR, Brownell KD (2009) Preliminary validation of the Yale Food Addiction Scale. Appetite 52: 430-436.
- Carlier N, Marshe VS, Cmorejova J, Davis C, M€uller DJ, et al. (2015) Genetic similarities between compulsive overeating and addiction phenotypes: a case for “food addiction”? Curr Psychiatry Rep 17: 96.
- Flint AJ, Gearhardt AN, Corbin WR, Brownell KD, Field AE, et al. (2014) Foodaddiction scale measurement in 2 cohorts of middle-aged and older women. Am J Clin Nutr 99: 578-586.
- Colditz GA, Hankinson SE (2005) The Nurses’ Health Study: lifestyle and health among women. Nat Rev Cancer 5: 388-396.
- Rockhill B, Willett WC, Hunter DJ, et al. (1998) Physical activity and breast cancer risk in a cohort of young women. J Natl Cancer Inst 90: 1155-1160.
- Gearhardt AN, Corbin WR, Brownell KD (2009) Food addiction: an examination of the diagnostic criteria for dependence. J Addict Med 3: 1-7.
- McAdams MA, Van Dam RM, Hu FB (2007) Comparison of self-reported and measured BMI as correlates of disease markers in US adults. Obesity (Silver Spring) 15: 188-196.
- Willer CJ, Li Y, Abecasis GR (2010) METAL: fast and efficient meta-analysis of genomewide association scans. Bioinformatics 26: 2190-2191.
- Ioannidis JP, Patsopoulos NA, Evangelou E (2007) Heterogeneity in meta-analyses of genome-wide association investigations. PLoS One 2: e841.
- Pe’er I, Yelensky R, Altshuler D, Daly MJ (2008) Estimation of the multiple testing burden for genomewide association studies of nearly all common variants. Genet Epidemiol 32: 381-385.
- Li MD, Burmeister M (2009) New insights into the genetics of addiction. Nat Rev Genet 10: 225-231.
- Liu JZ, McRae AF, Nyholt DR, et al (2010) A versatile gene-based test for genome-wide association studies. Am J Hum Genet 87: 139-145.
- Wu MC, Kraft P, Epstein MP, et al. (2010) Powerful SNP-set analysis for case-control genome-wide association studies. Am J Hum Genet 86: 929-942.
- Segre AV, Groop L, Mootha VK, Daly MJ, Altshuler D, et al. (2010) Common inherited variation in mitochondrial genes is not enriched for associations with type 2 diabetes or related glycemic traits. PLoS Genet 6: e1001058.
- McGue M, Zhang Y, Miller MB, et al. (2013) A genome-wide association study of behavioral disinhibition. Behav Genet 43: 363-373.
- Thorgeirsson TE, Gudbjartsson DF, Surakka I, et al. (2010) Sequence variants at CHRNB3- CHRNA6 and CYP2A6 affect smoking behavior. Nat Genet 42: 448-453.
- Genomes Project C, Abecasis GR, Auton A, et al. (2012) An integrated map of genetic variation from 1,092 human genomes. Nature 491: 56-65.
- Cuadrado A, Nebreda AR (2010) Mechanisms and functions of p38 MAPK signalling. Biochem J 429: 403-417.
- Chen RS, Deng TC, Garcia T, Sellers ZM, Best PM, et al. (2007) Calcium channel gamma subunits: a functionally diverse protein family. Cell Biochem Biophys 47: 178-186.
- Roadmap Epigenomics C, Kundaje A, Meuleman W, et al. (2015) Integrative analysis of 111 reference human epigenomes. Nature 518: 317-330.
- Ernst J, Kheradpour P, Mikkelsen TS, et al. (2011) Mapping and analysis of chromatin state dynamics in nine human cell types. Nature 473: 43-49.
- Murphy A, Tantisira KG, Soto-Quiros ME, et al. (2009) PRKCA: a positional candidate gene for body mass index and asthma. Am J Hum Genet 85: 87-96.
- GTEx Consortium (2013) The Genotype-Tissue Expression (GTEx) project. Nat Genet 45: 580-585.
- Hirose T, Apfel R, Pfahl M, Jetten AM (1995) The orphan receptor TAK1 acts as a repressor of RAR-, RXR- and T3R-mediated signaling pathways. Biochem Biophys Res Commun 211: 83-91.
- Liu CT, Monda KL, Taylor KC, et al. (2013) Genome-wide association of body fat distribution in African ancestry populations suggests new loci. PLoS Genet 9: e1003681.
- Fox CS, Liu Y, White CC, et al. (2012) Genome-wide association for abdominal subcutaneous and visceral adipose reveals a novel locus for visceral fat in women. PLoS Genet 8: e1002695.
- Hindorf L, MacArthur J, Morales J, et al. (2015) The NHGRI-EBI Catalog of published genome-wide association studies. Available at: https://www.ebi.ac.uk/gwas/.
- Pan Y, Wang KS, Aragam N (2011) NTM and NR3C2 polymorphisms influencing intelligence: family-based association studies. Prog Neuropsychopharmacol Biol Psychiatry 35: 154-160.
- Bajo M, Herman MA, Varodayan FP, et al. (2015) Role of the IL-1 receptor antagonist in ethanol-induced regulation of GABAergic transmission in the central amygdala. Brain Behav Immun 45: 189-197.
- Davis C, Loxton NJ, Levitan RD, Kaplan AS, Carter JC, et al. (2013) ’Food addiction’ and its association with a dopaminergic multilocus genetic profile. Physiol Behav 118: 63-69.
- Thorgeirsson TE, Gudbjartsson DF, Sulem P, et al. (2013) A common biological basis of obesity and nicotine addiction. Transl Psychiatry 3: e308.