A Novel Multimodal Strategy Combining Mirna Modulation and Tyrosine Kinase Inhibitors to Enhance Glioblastoma Cell Sensitivity to Therapeutics
Viraj Mehta1*
1 Basis Scottsdale High School, USA
*Corresponding Author: Viraj Mehta, Basis Scottsdale High School, USA, Tel: +1 480-451-7500; Fax: +1 480-451-7500; E-mail: viraj28m@gmail.com
Citation: (2020) A Novel Multimodal Strategy Combining Mirna Modulation and Tyrosine Kinase Inhibitors to Enhance Glioblastoma Cell Sensitivity to Therapeutics. Archiv Neurol Neurosurgery 3: 119.
Copyright: © 2020 Viraj Mehta, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: August 05, 2020; Accepted: September 16, 2020; Published: September 19, 2020.
Introduction
Glioblastoma Multiforme is the most common and lethal cancer of the adult brain, remaining incurable with a median survival time of only 15 months. Traditional efforts have focused on Receptor Tyrosine kinases (RTKs) such as epidermal growth factor receptor (EGFR) since EGFR is overexpressed in ~60% of primary glioblastomas and it activates downstream signaling that promotes tumor cell survival, proliferation and migration. The most frequently occurring EGFR mutation in glioblastoma, EGFRviii, arises from an in-frame deletion in the DNA sequence encoding the extracellular domain. EGFRviii is a cancer specific mutation and an attractive target for therapeutic intervention because it is not detected in normal tissues. Studies have indicated that EGFRviii is expressed in ~50% of glioblastomas that amplify wild- type EGFR.
Overexpression, amplification and mutation of EGFR is noted in many tumor cells, thereby supporting the notion that deregulated EGFR expression and signaling are pivotal events in the origin of human cancers. Hence, traditional efforts have focused on developing therapeutic agents and strategies to effectively inhibit EGFR. However, limited therapeutic benefits to cancer patients have been derived from EGFR-targeted therapies. One of the factors is blood brain barrier that limits the transport of drugs across to the central nervous system,while another factor is resistance to EGFR inhibiting drugs because of loss of phosphate and tensin homolog (PTEN). Research has shown that loss of PTEN is strongly correlated with treatment failure in GBM to the extent that the analysis of EGFRviii and PTEN levels may be used to predict tumor response to TKI (Tyrosine Kinase Inhibitors) basedtherapy.
One of the most comprehensive strategies aiming at identifying genes and pathways to mediate resistance to EGFR inhibition is the role miRNAs play in regulating these genes. The miRNA deregulation is involved in many processes, such as cell proliferation, cell cycle regulation, invasion, apoptosis, glioma stem cells behavior as well as angiogenesis. miRNA functions are significant in GBM oncogenic cell signaling and its potential to be considered as a biomarker and as a new therapeutic target for cancer.
Research Question: Can RTK inhibition in combination with the regulation of miRNAs through small molecules hold promise as a prospective strategy towards GBM treatment vs standard therapy of using commercial Temozolomide? Hence, the hypothesis of this research project lies in the statement that regulating miRNAs with Small molecules as inhibitors along with TKIs is a better approach towards controlling glioma progression compared to targeting glioma cells with commercially available anticancer drugs(TMZ). The high-level strategic view is shown in the figure below: [1].
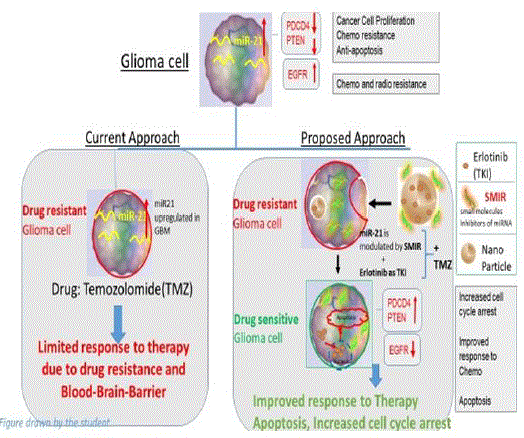
Methods and Materials
In order to find key proteins that are overexpressed, mutated or deleted, STRING software was used to create protein-protein interaction maps and focus on the most critical proteins with maximum interactions (Figure 1).
Figure 1: Protein-Protein Interactions using STRING software [2].
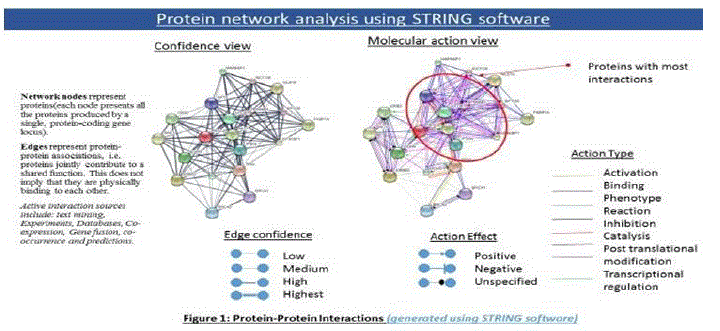
The PPI interactions show TP53, PTEN, EGFR, KRAS, mTOR as the proteins with most interactions, and initiating either binding, inhibition, post translational modification, or activation. In order to correlate this with actual data, TCGA (The Cancer Genome Atlas) database for patient sample data was used (Figure 2) which showed PTEN as the protein which was deleted in most cases (36%) while EGFR was the receptor amplified or overexpressed in most cases (53%).
Figure 2: TCGA patient sample data [3].
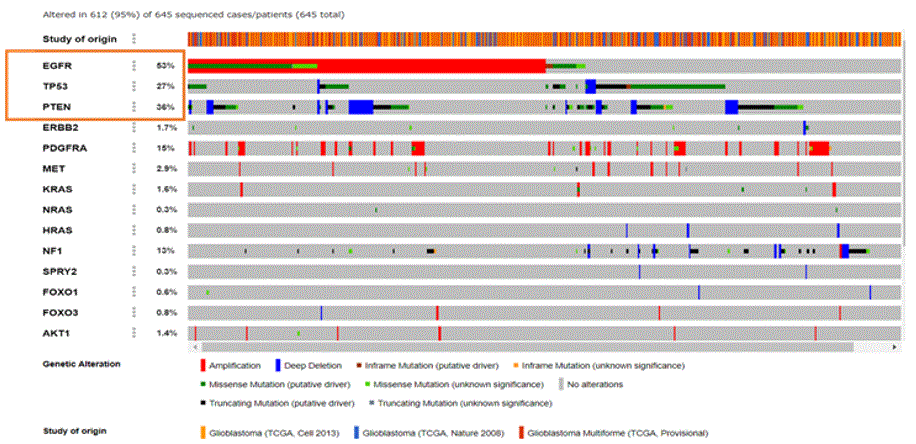
As seen from the TCGA data, EGFR was overexpressed in 53% of the samples and PTEN gene deletion was observed in 36% cases, whereas tumor protein TP53 mutations (missense & truncating) were found in 27% of the samples. Amplification of EGFR is frequently associated with the occurrence of a mutant form of EGFR called EGFR variant III (EGFRviii, also known as ΔEGFR). EGFRviii is found in approximately 50% of all glioblastomas [16]. An overview of Glioblastoma pathways was downloaded from Kanehisa labs [2] and is shown in Figure 3(a) below.
Figure3a: Overview of GBM Pathways.
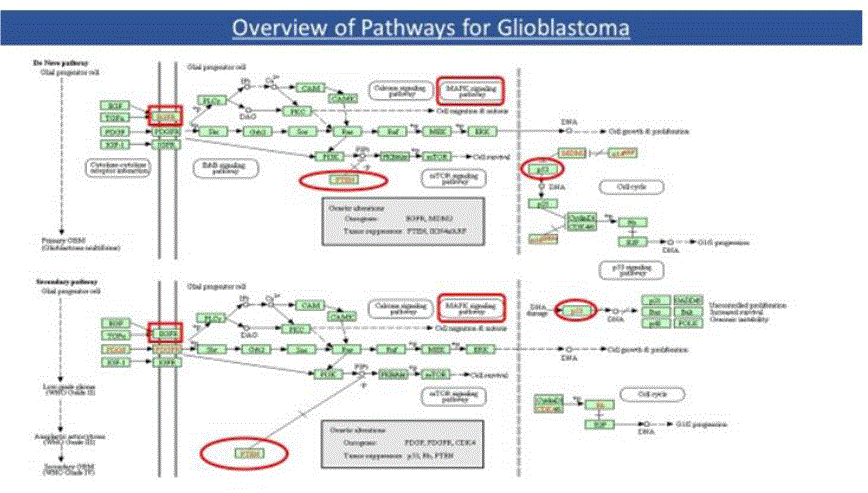
From the above, it can be seen that the Glioma relevant activation points of the Ras/Raf/MAPK(ERK) pathway is via amplification(or mutation) of RTKs, such as the EGFR/PDGFR, and of the growth factor receptor-PI3K pathway by loss of the PTEN tumor suppressor gene. In addition, Primary glioblastomas develop in older patients and typically show alterations such as EGFR amplification, p16/INK4a deletion, and PTEN mutations at frequencies of 24-34%. Secondary glioblastomas develop in younger patients and frequently show overexpression of PDGF and CDK4 as well as p53 mutations (65%) and loss of Rb. Loss of PTEN is much more common in the pathogenesis of primary GBM.
A simplistic view of PI (3) K and MAPK pathways was drawn as shown below in figure 3(b):
Figure 3b: PI (3) K and MAPK Pathways.
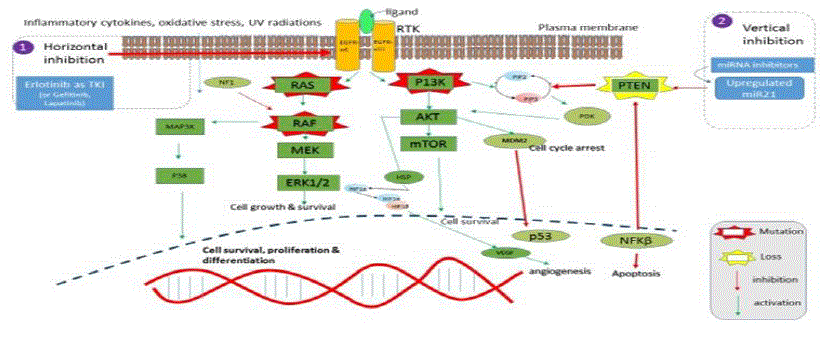
The activation of PI3K starts by the binding of a ligand to a receptor tyrosine kinase (RTK). Receptor tyrosine kinases (RTKs) are a family of cell surface receptors, which act as receptors for growth factors, hormones, cytokines, neurotrophic factors and other extracellular signaling molecules. Upon activation by ligands, RTKs signal through two major downstream pathways Ras/MAPK/ERK and Ras/PI3K/AKT. These pathways are involved in the regulation of cell proliferation, survival, differentiation and angiogenesis, and hence are promising therapeutic targets for the treatment ofGBM. The RTK activation by phosphorylation of its intracellular domain activates the catalytic subunit of PI3K and this results in the generation of PIP3 from PIP2. PIP3 in turn phosphorylates AKT. AKT phosphorylates many downstream targets to induce NFκB activation and mTORC2 and MDM2 to induce cell survival. PTEN, a tumor suppressor gene antagonizes the PI3K pathway by dephosphorylating the second messenger PIP3 to PIP2. PTEN loss promotes resistance to EGFR by dissociating EGFR inhibition from downstream inhibition of the PI3K pathway. There is extensive data suggesting that the loss of PTEN by mutation, methylation, or deletion leads to a decreased apoptosis and that the loss of PTEN is frequent in GBM. Hence, this study focused on EGFR and PTEN.
Studies indicate that miRNAs regulation plays a significant role in glioma development and Glioblastoma pathogenesis. Its role in cell proliferation, apoptosis, cell cycle regulation, migration, invasion, and angiogenesis is critical and defects in miRNA regulatory network appear to play a key role in glioblastoma pathogenesis. Hence, DIANA-Mir Path computational tool was used to map microRNAs to proteins (KEGG pathways) they regulate during various stages of Glioma progression. It was found that up-regulated miR-21 leads to reduced apoptosis by modulating TP53, and initiates tumorigenesis by downregulating PTEN and PDCD4 (tumor suppressor genes). See Figure 4a below.
Figure 4a: miRNA mapping to Oncogenes/Tumor suppressors [4].
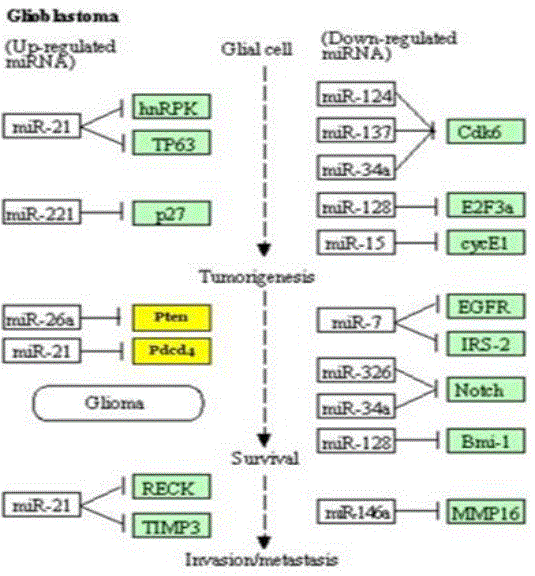
Figure 4b: An overexpressed or upregulated miR21 inhibiting PTEN.
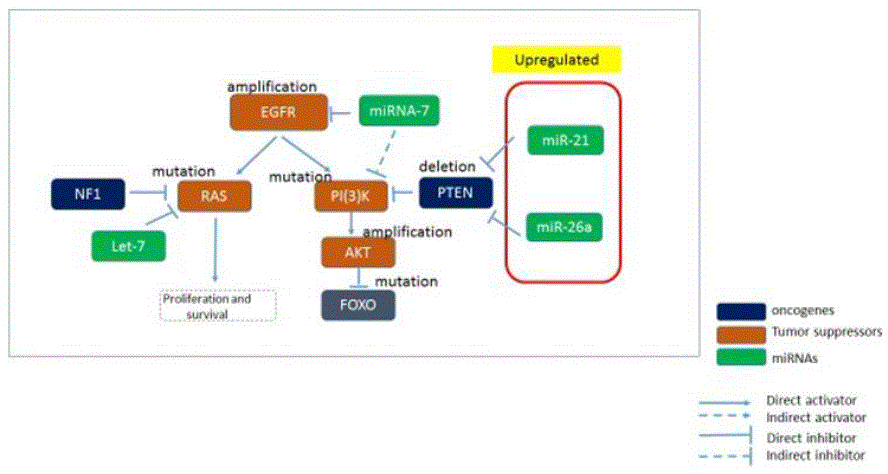
While miR21 and miR26 are upregulated and can be inhibited by small molecules with inhibiting properties, there are down-regulated miRNAs, e.g. miR-7 which causes EGFR overexpression. MiR-7 is a regulator of the phosphoinositide-3-kinase (PI3K)/ATK and Raf/mitogen-activated protein kinase (MEK)/extracellular signal-regulated kinase (ERK) pathways, both of which are launched by EGFR through its two direct targets, the transcription factors PI3K and Raf-1, respectively. Enforcing miR7 expression has been related to a marked decrease expression of P13K, AKT, RAF-1, as well as of EGFR.
It is reasonable to conclude that miR-21 is consistently upregulated in GBM and targets numerous pathways including those involved in survival, proliferation, invasion, and apoptosis. This is also substantiated by experimental research that miR-21 is a well-known oncomiR and is involved in all steps of tumor development, including initiation, maintenance and survival.
In order to analyze the interactions of miRNAs with mRNAs, DIANA tools (mirExTra2.0) was used to develop the differential analysis on miRNAs, and explore the up/down regulated miRNAs vs mRNAs. The DIANA-mirExTra v2.0 web server utilizes miRNA/mRNA, TF/mRNA and TF/miRNA interactions derived from extensive experimental data sets. miR-21 was found to have highest prediction score (0.999) for PTEN, hence was chosen for further analysis for its druggability with small molecules [5].
Molecular Docking between molecules – Background
The basis of all processes in living organisms lies in molecular recognition where macromolecules, such as proteins interact with other small molecules to form a specific complex. Protein-ligand interactions are hence foundational to understanding biology at the molecular level. In addition, the underlying mechanisms for the protein-ligand recognition, affinity and binding facilitate drug discovery, design and development. Molecular docking is a tool to predict the predominant binding mode between a ligand and a protein of 3D structure, and is primarily used for structure-based drug design or discovery [6].
It allows exploring the binding and best fit between a small molecule and its receptor and hence enable one to derive a conclusion on the druggability of the receptor molecule. The protein- ligand docking methods helps predict the correct bound conformations in a more time and cost- effective manner, making it very suitable for high-throughput virtual drug screening. Essentially, Docking predicts the best binding modes between the 3D structures of two molecules by fitting the ligand into the pocket and maximizing its affinity. The two steps involved in a successful docking include pose generation and assignment of a scoring/numerical value to the molecule.
- Scoring functions predict the energy of a particularpose.
- Search methods find the optimal pose.
Scoring function, measured by the binding affinity, is calculated by the formula:
δGbinding = δGvdW + δGelec + δGHbond + δGdesolv + δGtors where the terms are defined as below: δGvdW (Van der Walls dispersion repulsion): models the shape of the ligand with its complimentary receptor.
δGelec (electrostatic term): interaction between phosphate & metal ion
δGHbond: interaction between base and carboxyl group.
δGdesolv: entropic and enthalpic contributions of release of water molecules that surround ligand and fill the ligand when the binding is done, models hydrophobicity effect.
δGtors: accounts for entropy of freezing out all degrees of freedom. If ligand is very flexible, this is an unfavorable term [7].
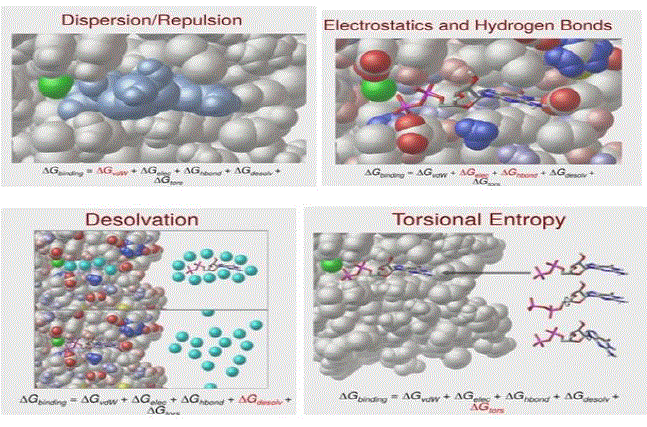
The driving forces behind the association between protein and ligands are a result of various interactions and energy exchanges among the proteins, ligands, water molecules, and buffer ions. Protein–ligand binding occurs only when the magnitude of binding energy is high, and because the protein–ligand association extent is determined by the magnitude of the negative δG, it can be inferred that δG determines the stability of a given protein–ligand complex, and hence the binding affinity of a ligand to a given acceptor. This is the basis of computational drug design and the approach to accurately predict the affinity of a drug to its target receptor. The affinity optimization of drug candidates is a key component of drug development, hence in order to enhance the binding affinity, the binding free energy between the lead compound and the target protein needs to be minimized through enthalpic and/or entropic contribution optimization.
In this case, 2 scenarios were explored:
Control case: EGFR Docking with TMZ
Preparation of Ligand: Ligand is an ion or molecule that binds to a protein site. UCSF Chimera, a highly extensible program for molecular modeling and visualization was used as the GUI for all simulations and structural editing. TMZ was downloaded from PubChem in its Canonical SMILES format: CN1C(=O) N2C=NC(=C2N=N1)C(=O)N
As a note, SMILES (simplified molecular-input line-entry system) is a specification in form of a line notation for describing the structure of chemical species using short ASCII strings. This structure was minimized by adding hydrogens and partial charges, and saved in .mol formatto get it ready fordocking.
Preparation of Receptor: Crystal structure of EGFR kinase domain 5U8L was downloaded from RCSB Protein databank site, in .pdb format into Chimera window and native ligands like sulphate ion, Glycerol etc. form chain A as well as solvents, water molecule were removed from the receptor molecule. Hydrogen atoms and charges were added back to the receptor, and the structure was saved in mol2 format. The hydrogen atoms and partial charges were added to the receptor to avoid any single residual atom as it may interfere with the docking being available to accept or donate electric charge to any other atom. The non-standard residues constituting ligand were removed before docking in order to leave the binding pocket empty because otherwise, the ligand could not bind within the binding site. The ions were removed to avoid any interference with the available charge as they may form additional bonds with the ligands.
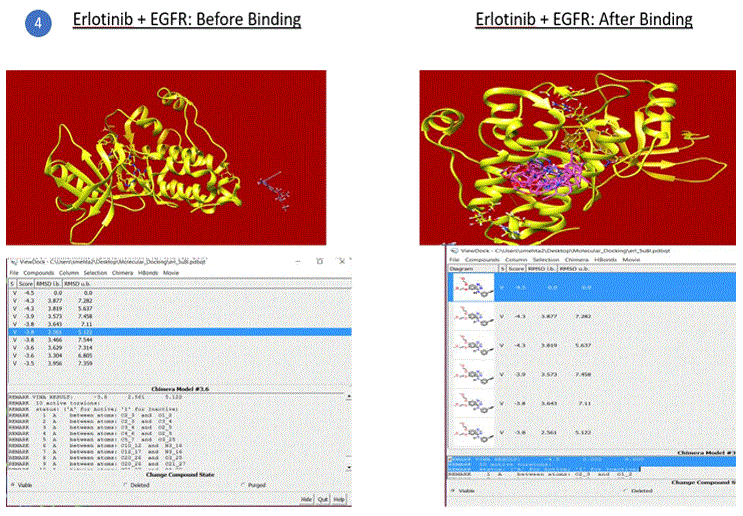
Preparation of AutoDock environment for simulations: Autoligand was used to predict the binding site of the receptor. AutoDock Vina was launched and a 3D grid site was defined where the binding must occur between the receptor and ligand. Receptor and Ligand options were defined (ignore water, ignore nonstandard residues, add hydrogens etc.). Simulation run was completed for EGFR+TMZ which produced an output file with a number of poses, the energy scores of the viable structures, RMSD (root mean square deviation) lower bound and upper bound values. The lowest energy value with 0 RMSD is the best conformation and was used to determine if the binding and hence the required outcome (e.g. inhibition in this case) was successful relative to other combinations. The best pose, shown in figure 5, is chosen on the basis of the conformation of the ligand in the binding site of the protein, i.e. how properly a ligand can fit in the binding pocket, which is typically fulfilled by the first or the second pose.
Figure 5: Final Complex (bound) structure of EGFR + TMZ, drawn using AutoDock S/W and Chimera GUI.
Test case: miR21 docking with Small Molecule Inhibitors and EGFR+Erlotinib
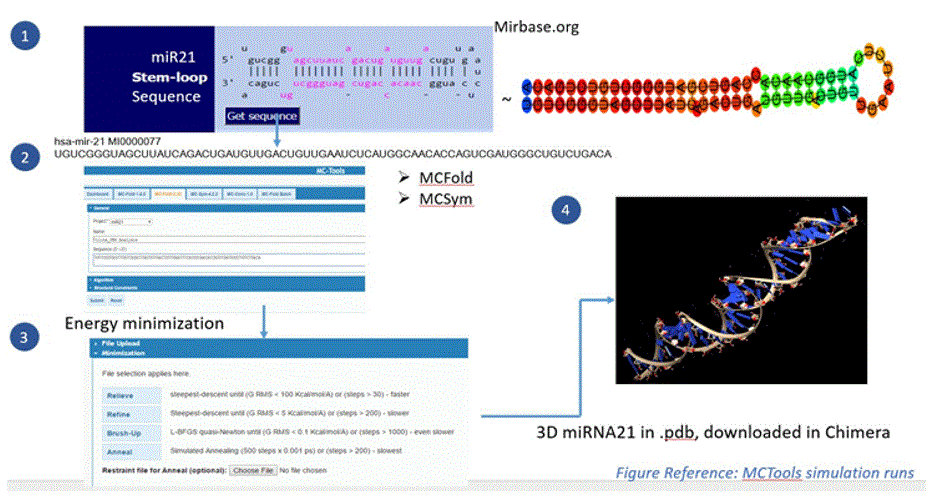
Preparation of miR21: Since miRNA driven strategies are fairly new in the field of cancer, RNA structures are not readily available like protein structures are in Protein databank (PDB). Therefore, the miR21 3D structure was created through the following process:
-Stem loop sequence was generated using miRbase software [9].
-This sequence was given as an input into MCFold2.32/MCSym pipeline.
-The MC-Fold/MC-Sym pipeline is a web-hosted service for RNA secondary and tertiary structure prediction. MCTools offers a suite of programs to analyze, model, predict, and determine RNA 2D and 3D structures.
-The pipeline consists of uploading miRNA sequence to MC-Fold, which outputs (2D) secondary structures that are directly input to MC-Sym, MC-Sym then outputs the (3D) tertiary structures [10].
In the current research, MCFold 2.32 created 50 2Dstructures with energy values ranging between -76kcal/mol to -78.92kcal/mol. First 10 values based on lowest energy (highest absolute value) were chosen to be given as an input into MCSym pipeline to create a stable 3D structure.
Energy optimization and minimization was further conducted through the “Relieve- Refine_Brushup-Anneal” process of MCTools.
Minimum Free Energy as one of the deterministic methods is used by MCTools based on the hypothesis that an RNA molecule will fold into a secondary structure that minimizes its free energy
W(i, j) =0, if i = j ; min{V (i, j), min i≤k≤j{W(i, k) + W(k + 1, j)}},
Final 3D miR21 structure was saved in the pdb format and imported into the Chimera GUI for docking with small molecules. The miR-21 3D structure preparation is shown in the figure below:
Figure 6: miR21 3D structure preparation (using mirbase.org and MCTools).
Preparation of small molecules as Ligands: 1. 500 chemical compounds were taken from PubChem [11] and downloaded in the “Canonical SMILES” format into the Chimera GUI for autodocking against the 3D miR-21.
Molecular docking and post docking analysis: High-throughput Docking-based virtual screening was conducted using AutoDock program version 4. The grid based binding sites were defined, protein-ligand interactions were analyzed and visualized using Chimera. The energy scoring of all docking simulations is given in Table 1.
Table 1: Simulation results of the test case and the control case.

Simulation results of docking are also shown in the pictures below. Note: Following pictures are results of AutoDock simulation runs by the student using Chimera as a GUI
Results and Discussion
The control case and the test case & the Results of simulations are tabulated below: 1. EGFR +TMZ. 2.miR21+ Small molecules as inhibitors (AC1MMYR2, AC1L1BBW.)+Erlotinib.
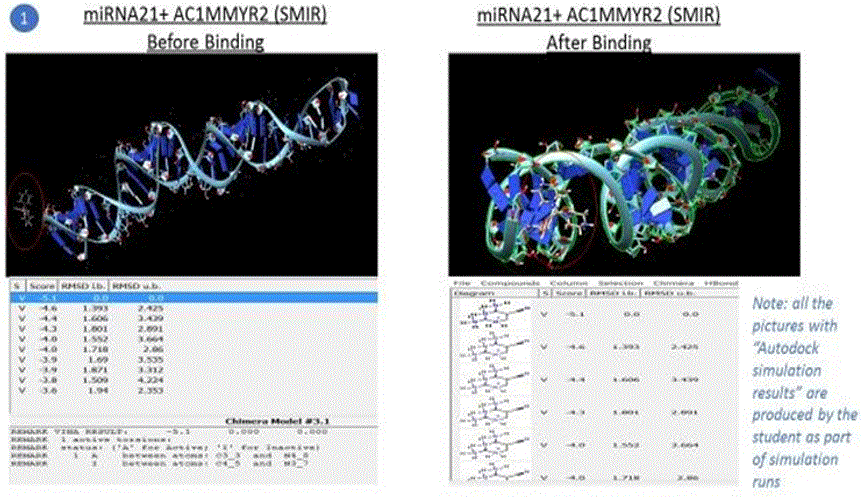
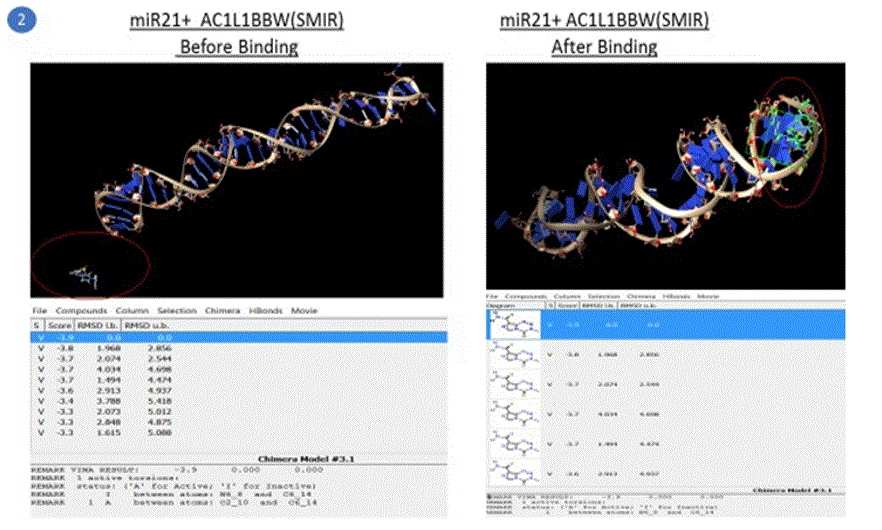
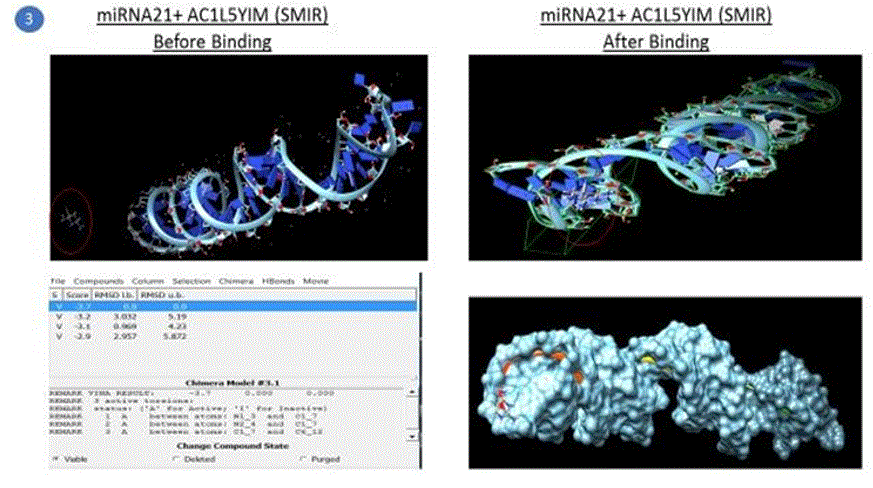
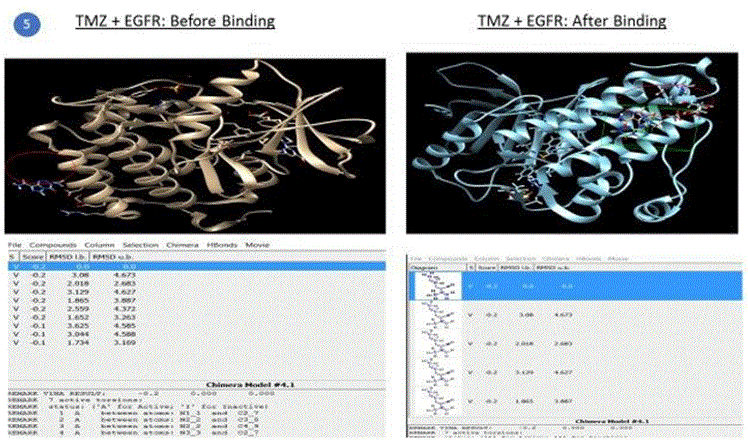
Five compounds out of the 500 compounds I ran the simulations for were found to be selective inhibitors of miR21 based on their highest binding scores (lowest energy in kcal/mol) as shown in the results table1 above. These were compared against the docking results of EGFR with TMZ and EGFR + Curcumin (diferuloylmethane, known to have therapeutic potential against cancer), also shown in the table 1 above.
The binding energy of the complex EGFR + TMZ was found to be relatively high at - 0.2kcal/mol compared to -5.1kcal/mol for miR21+AC1MMYR2 (small molecule as inhibitor). It is worth to note that AC1MMYR2 and miR21 complex gave 10 poses, with only 1 active torsion. Note that a pose is a binding mode and number of torsion angles decide ligand flexibility for docking. The highest binding affinity and the corresponding absolute free energy at -5.1kcal/mol was considered the zero mode with zero RMSD upper and lower bounds. On the contrary, EGFR+TMZ complex had a relatively high binding energy at -0.2kcal/mol, and gave 7 torsions with 10 poses. (Torsions should be minimal for a stable compound). EGFR+CUR complex gave 3 torsions, and 10 poses but a low absolute energy of -0.9kcal/mol. In contrast, all the 5 SMIR+miR21 combination outputs gave lowest energy compounds ranging between - 2.8kcal/mol to -5.1kcal/mol. (note that as the free energy of the system decreases, the system becomes more stable i.e A low (negative) energy indicates a stable system and thus a likely binding interaction).
This confirms the hypothesis that regulating miRNAs with small molecules to modulate tumor suppressor gene (PTEN) expressions followed by sensitizing glioma cells with TKIs is a better approach to improving drug sensitivity rather than targeting protein receptors directly with therapeutic drugs. The results of AutoDock simulations demonstrated small molecule AC1MMYR2 had the highest binding affinity to miR21, and hence a strong anti-tumor drug candidate.
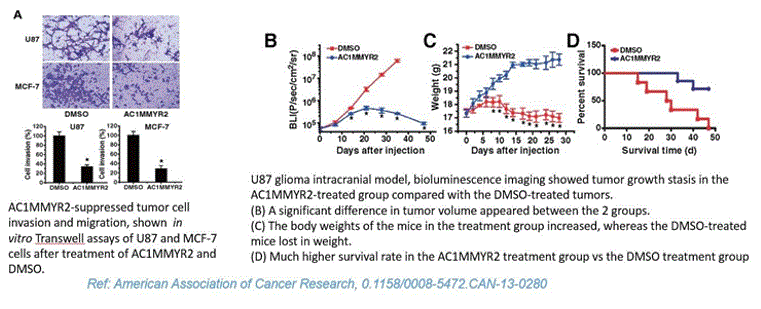
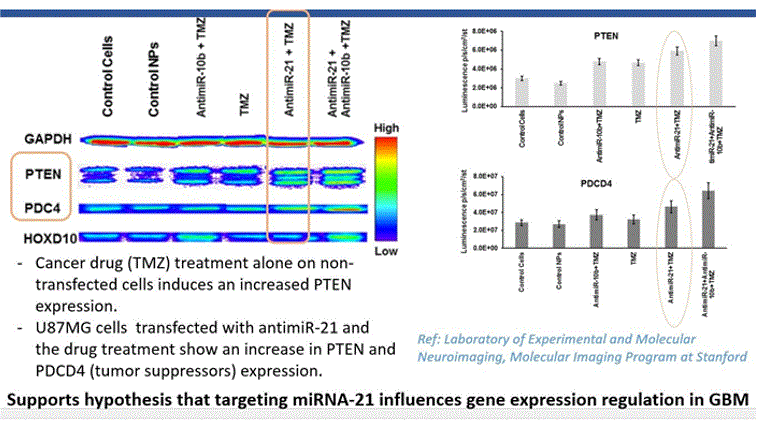
The above conclusion based on docking simulations were substantiated with the immunoblot analysis of U87MG cells transfected with antimiR-21, which showed an increase in PTEN and PDCD4 (tumor suppressors). Additionally, lab results from American Association of Cancer Research showed significant decrease in tumor volume after treatment with AC1MMYR2, an inhibitor of miRNA21 (Results shown below)
Analysis
Cancer cells have a fundamental characteristic of being able to continuously proliferate, primarily due to deregulation of cell signaling pathways. The phosphatidylinositol-3-kinase (PI3K)/AKT and MAPK signaling pathway are crucial to many aspects of cell growth and survival. The recognition of its importance in tumorigenesis and cancer progression has led to the development of a number of agents that target various components of these pathways as cancer therapeutics. However, the therapeutic efficacy of single-agent PI3K pathway inhibitors is likely limited by feedback regulations among its pathway components and crosstalk with other signalingpathways.
In addition, this promotion of sustained signaling for proliferation is influenced by microRNAs, which also evade tumor growth suppressors, further enhancing cancer cell growth. For example, miR21 is frequently overexpressed in GBM, and hence downregulates tumor suppressors PDCD4 and PTEN, which then promotes cell cycle progression, tumor invasion, metastasis and resistance to therapeutic drugs. Overexpression of miR-21 significantly inhibits the effect of TMZ on apoptosis and studies have shown that by inhibiting an upregulated miR-21, chemo sensitivity of human GBM cells can be enhanced. The current research and data collected using computational analysis by the author, further sets out to prove that miRNA modulation in combination with TKIs can dramatically improve the susceptibility of glioma cells to MTKIs like Erlotinib, Sunitib etc. whereas no significant improvement is observed when glioma cells are treated with TMZ directly without prior reprogramming of cells with TKIs or miRNA regulated.
The Cancer Genome Atlas (TCGA) data shows that the Primary GBM is frequently associated with epidermal growth factor receptor (EGFR) amplifications. Aberrant amplification, deletion, or mutation of at least one receptor tyrosine kinase (RTK) has been found in 67% of all GBM, with EGFR accounting for 53% cases (Figure 3). It is generally accepted that key features of GBM are not only the reflection of dysregulated pathways and associated protein functions but also the consequence of the dysregulation of miRNA-mediated translation control [9]. Whether a miRNA acts as an oncogene or a tumor suppressor depends on the regulated genes and cellular context. The impact of miRNAs on the PI3K/MAPK pathways was analyzed because over or under expression of miRNA has been linked to dysregulated pathways. The cancer genome atlas (TCGA) data portal was used to obtain miRNA expression profiles of 645 cases with GBM. PTEN gene deletion was observed in 36% of cases and EGFR was overexpressed in 53% of the patient samples. This research focused on PTEN tumor suppressor and miRNAs, which influence the regulation of PTEN. Through the miRbase and miRPath analytical tools, miR21 was found to inhibit PTEN and PDCD4. Research on glioma cancer cells has shown that when miR21 is inhibited, PTEN expression increases, thereby decreasing tumor cell proliferation and invasion. Mutant PTEN and EGFRviii are also negative regulators of the P13K/AKT signaling pathway, the activation of which is associated with reduced sensitivity to cancer drugs likeTemozolomide.
This research focused on a combinational approach of targeting Tyrosine Kinases with inhibitors to regulate overexpressed EGFR as well as modulating upregulated miRNAs through small molecule inhibitors. Specifically, AutoDock based virtual screening was used to find small molecule inhibitors of miR21 in order to regulate the PTEN expression. Docking as a structure- based drug discovery approach was used to analyze the binding between 3D protein and small molecule structures. DIANA-miRPath and microT algorithms were also used to predict interactions between tumor suppressors/oncogenes with drugs and upregulated miRNA with inhibitors with a certain prediction score.
The results of five compounds out of the 500 compounds I ran the simulations for, found to be potent and selective inhibitors of miR21 based on their highest binding scores (lowest energy in kcal/mol), were compared against the docking results of EGFR with Temozolomide (commercial anti-cancer drug). The binding energy of the complex EGFR + Temozolomide was found to be -0.2kcal/mol compared to -5kcal/mol for miR21+SMIR and -4kcal/mol for EGFR + Erlotinob. This demonstrated that combinational targeting of miRNAs with small molecules (miR21+AC1MMYR2 in this case) and protein receptors with TKIs(EGFR + Erlotinib) is a better approach to regulating gene expressions and controlling metastasis than targeting protein receptors directly with drugs(EGFR+TMZ). This was confirmed through the immunoblot analysis of U87MG cells transfected with antimiR-21(same as miRNA inhibitor) followed by Temozolomide drug, which showed an increase in PTEN and PDCD4 (tumor suppressors). The in-vivo experiment (reference: American association of cancer research) demonstrated that small molecule inhibitors of miR21 can block the ability of Dicer to process pre-miR-21 to mature miR-21, thereby suppressing tumor growth, invasiveness and metastasis. An increase in the PTEN(tumor suppressor) expression is observed after an upregulated miR21 is transfected with its inhibitor which reprograms the glioma cells, and increases their chemo-sensitivity by inhibiting the transition from G2/M phase to G1 phase of the cell cycle. In addition, miR-21 downregulation increases PDCD4 expression in Glioblastoma tumor cells resulting in decreased proliferation, increased apoptosis, and decreased colony formation[11]. The susceptibility of glioma cells to multitargeted TKIs such as Erlotinib (or Sunitinib) is also much higher after the miRNA modulation.
Hence a novel multimodal strategy combining Tyrosine kinase inhibitors (TKIs) and microRNA (miRNA) modulation was studied to overcome the signaling pathway deregulation and computationally prove a way to promote tumor cell death in case of Glioblastoma.
Conclusion
The critical pathways controlling cell growth and proliferation during cell development are the PI3K-AKT and MAPK pathways. PI3K gets activated in response to cell surface tyrosine kinase receptor-ligand binding, and subsequently converts PIP2 through phosphorylation to PIP3. The conversion of PIP2 to PIP3 can be reversed by PTEN, which therefore serves as a negative regulator of PI3K activity. However highly upregulated microRNAs can inhibit the tumor suppressor activity of PTEN. This study focuses on complex interactions of miRNAs, genes, Kinases and small molecules. The small molecules can bind to miRNAs (structure based binding approach) and regulate gene expressions such that the genes can be silenced/programmed to their normal function (e.g. PTEN as tumor suppressor) before treating with anti-cancer drugs.
Given the therapeutic necessity of simultaneously targeting multiple pathways in Glioblastoma, the ability of individual miRNAs to target multiple genes/pathways and modulate key cellular processes is a major advantage, making miRNAs ideal therapeutic targets for Glioblastoma. DIANA-miRPath tool identified microRNA-21 (miR-21) as one of the key oncomiRs that is significantly overexpressed in glioblastoma, controlling its tumor invasion, apoptosis, cell proliferation, and chemo resistance. miRNA21 is also the most prevalent microRNA impacting multiple pathways and proteins including P13K/AKT, RTK/RAS, PTEN, and PDCD4 and is found to be significantly upregulated in Glioblastoma, thus repressing the tumor suppressor PTEN pathway and advancing tumor progression (Figure 4a).
Small molecules from PubChem database were virtually screened using AutoDock software to select the ones that can block miR-21 maturation. AC1MMYR2 small molecule was identified to most effectively bind with miR21 with a binding energy of -5.1kcal/mol, and block miR-21 maturation, regulating PTEN expression thereby suppressing tumor growth and progression. Thus, the simulations and data analysis predicted the druggability of microRNAs, and demonstrated the high binding affinity of an upregulated miR-21 with its inhibitor AC1MMYR2. The results were validated with the lab experiments that show AC1MMYR2 blocks the ability of Dicer to process pre-miR-21 to mature miR-21 and upregulates the expressions of PTEN and PDCD4 thereby suppressing proliferation, survival, and invasion in glioblastoma[10]. miRNA-21 modulation with its inhibitor, hence promoted the most pronounced sensitization of GBM cells to TKI, specifically Erlotinib.
This research proves that small molecules with structural similarities to miRNAs canbind through a structure-specific interaction and provide an opportunity to improve the responses of Glioma cells to drugs and chemotherapy. The novelty of this approach lies in the fact that instead of directly targeting receptors or genes (EGFR, PTEN) that tend to be insensitive to drugs and chemotherapy, it is more effective to regulate the receptors or genes (EGFR, PTEN) through miRNAs, which can easily bind with small molecules and enable gene silencing thereby controlling the tumor progression. This vertical inhibition when combined with horizontal inhibition of pathways using TKIs like Erlotinib can produce a synergistic effect on regulating the PI (3)K and MAPK pathways. Additional benefit is that this approach can be used to screen drugs for any kind of cancer. The premise of this research is based on SBDD (Structure based drug discovery) where known protein structure is used to bind with the “ligand binding” small molecule compounds and the binding affinity of the complex is used to decide the strength of the interaction between thetwo. This computer-aided drug discovery approach provides a promising tool to discover a solution to regulating cancer cell proliferation in a more time and cost-effective manner, and provide further insight into traditional lab-based investigations.
Future Research
The above research was done using structural similarities between drugs and proteins. It will be a logical next step to understand the role of MORFs (molecular recognition features) - short regions of around 20 residues that intrinsically disordered proteins (IDPs) contain. MORFs undergo a disorder to order transformation when IDPs bind to other proteins. I would like to explore ways to inhibit IDP’s protein-protein interactions by designing drugs that can bind to IDP MORFs or design small molecule drugs resembling IDP’s MORFs by leveraging the structural similarity of small molecule drugs to IDP’s binding site.
Another future step would be to use in-vivo/in-vitro methods to investigate multiple SMIR and miRNA combinations. Additionally, using data from patients’ unique genomes and discovering upregulated vs downregulated miRNAs (e.g. miR-7) that are influencing the pathways to develop personalized treatments will contribute to more effective therapies for GBM. Targeted drug delivery has been an ongoing challenge for GBM treatment, mainly because the blood brain barrier acts as a diffusion barrier and impedes the influx of compounds including anti-cancer drugs from blood to the brain. I also would like to investigate the improvement in drug delivery with variations in nanoparticles and sequential co-delivery of drugs such as TMZ/DOX and miRNAs.
References
- Author’s hypothesis.
- https://string-db.org/
- http://www.cbioportal.org/
- http://snf-515788.vm.okeanos.grnet.gr/index.php?r=mirpath/pathwayviewer
- http://diana.imis.athena- innovation.gr/DianaTools/index.php
- https://www.rcsb.org/pdb/explore/explore.do?structureId
- http://www.ncbi.nlm.nih.gov/sites/entrez?cmd=search&db=pccompound
- http://www.ncbi.nlm.nih.gov/sites/entrez?cmd=search&db=pccompound